
В первых трех частях данной серии (часть 1, часть 2, часть 3) были рассмотрены многочисленные данные по гаплогруппам и субкладам/снипам ископаемых ДНК, опубликованные в первой половине 2017 года. Эти данные были в значительной степени суммированы в двух протяженных таблицах, опубликованных в приложениях к работам Mathieson и др. (2017) и Olalde и др. (2017), и содержащих 469 и 480 данных по ископаемым гаплогруппам и субкладам, соответственно (268 и 277 Y-хромосомных данных, соответственно, остальные – только мтДНК), с территориями и археологическими датировками. Помимо того, в 2017 году были опубликованы многие десятки данных по ископаемым гаплогруппам и субкладам в статьях других авторов, которые были по возможности приведены в трех предыдущих статьях данной серии. Это, в совокупности с более ранними таблицами, опубликованными в 2015 году в статьях Allentoft и др. (230 ископаемых гаплогрупп и субкладов, из которых 126 – Y-хромосомных) и Haak и др. (69 Y-хромосомных ископаемых гаплогрупп и субкладов), предоставили исключительно важную основу для анализа древних миграций. Большую и важную работу в этом отношении провел И.Л. Рожанский, который нанес эти и другие многочисленные данные на 12 карт, организованных по историческим периодам, с времен ранее 11 тысяч лет назад и до нашего времени (последнее – за исключением большинства стран Восточной Европы).

Лепенский Вир (современная реконструкция древнего поселения)
Таким образом, появилась возможность критически рассмотреть более ранние концепции о древних передвижениях родов и племен, условно принимая гаплогруппы как рода, и субклады как племена, хотя границы между ними часто размыты – например, гаплогруппа R1b – это технически, да и по сути дела, субклад, а гаплогруппа – R, хотя и она технически может рассматриваться как субклад гаплогруппы Р. Поэтому при подобном рассмотрении важен контекст, в котором используются понятия гаплогруппы, субклада, рода, племени. Не лишне в очередной раз напомнить, что ДНК-генеалогия не рассматривает этносы, национальности, партийности и тому подобное, хотя рода входят в качестве составных частей в каждое из этих понятий. Уже совершенно ясно, что при изучении этногенеза необходимо рассматривать рода, как их понимает ДНК-генеалогия, и что этногенез в значительной степени определяется динамикой взаимодействия родов, при дополнительном влиянии многих исторических, военных, культурных и других факторов.
Под «более ранними концепциями» о древних передвижениях носителей гаплогрупп и субкладов здесь подразумеваются концепции, выработанные – как правило, в предварительном виде – на основании современного распределения гаплогрупп и субкладов, расчетов хронологии, динамики их развития, на основании картины мутаций в перекрестных выборках гаплотипов их представителей. Просто описательное распределение гаплогрупп и субкладов по регионам мало о чем говорит в отношении истории их развития. О чем, например, говорит тот факт, что у басков наблюдается 85% гаплогруппы R1b? Пожалуй, только о том, что у них доминирует гаплогруппа R1b. Про историю басков это практически ничего не говорит. А расчеты по мутациям в выборках гаплотипов современных басков этой доминирующей гаплогруппы показывают, что их общий предок жил примерно 4900 лет назад (Клёсов, «История ариев и эрбинов», 2017, стр. 279-280). Расчеты без учета ветвей субкладов, «поперек» их доминирующей гаплогруппы R1b-P312, дают датировку 3625±370 лет назад (Klyosov, 2009). Кстати, недавно те же R1b-P312 найдены в захоронении в Португалии с археологической датировкой 3740-3430 лет назад (Olalde et al., 2017). Так что здесь палеогенетические данные вовсе не противоречат тем, которые были расчитаны по гаплотипам современников (см. здесь).
Но популяционные генетики традиционно придерживаются примитивных представлений – если где-то наблюдается высокая доля определенной гаплогруппы, значит, они там жили всегда, или во всяком случае являются «коренным народом», автохтонами. Более того, если процент гаплогруппы самый высокий – то она там, по их мнению, и образовалась. Типичные примеры – ранние представления о том, что гаплогруппа R1b образовалась в Европе, при этом бралась «потолочная» цифра, что образовалась там 30 тысяч лет назад. Основания – потому что гаплогруппы R1b в Европе «больше всех». А 30 тысяч лет – потому что «еще при неандертальцах», «вот какие мы древние».
Еще пример – в статьях индийских ученых совместно с российскими и эстонскими попгенетиками серьезно утверждалось, что поскольку среди брахманов высшей касты в Индии содержание гаплогруппы R1a «самое высокое в мире», то гаплогруппа R1a якобы образовалась в Индии. На это можно возразить тем, что в нашей фамильной деревне в Курской области содержание R1a равно 100%, а значит, гаплогруппа R1a образовалась в деревне Клёсово, или во всяком случае в Курской области России. Но ссылки на те фантасмагорические статьи индийских, эстонских и российских попгенетиков до сих пор висят на «моей» страничке в Википедии (как якобы доказательство «лженаучности ДНК-генеалогии», как и ссылка на не менее фантасмагорическую статью Л. Клейна, который, согласно ссылке, рассуждал о том, что она (гаплогруппа R1a1) выделилась из корня R1a в Индии в период мезолита 14 тысяч лет назад [30] [31], задолго до вторжения ариев в Индию и Иран». Фантасмагорическая – потому что Клейн, ничего не понимая в ДНК-генеалогии, списал эту «датировку» у попгенетиков, которые использовали для «расчетов» печально известную «популяционную скорость Животовского», от которой сами попгенетики уже отказались, а Клейн блистает с ней на страничке в Википедии. Ссылка [30] – это его, Клейна, а [31] – скопом на серию статей попгенетиков со столь же безумными выводами, что и у Клейна, у которых он, Клейн, банально списал.
Еще пример – на Балканах содержание гаплогруппы I2а в целом самое высокое в мире, например, в Боснии-Герцеговине 51% от всего населения, среди боснийских хорват оно составляет 71%, и попгенетики традиционно утверждали, что I2а там, на Балканах, и образовалась. Однако ДНК-генеалогия уже давно, 8 лет назад, показала, что общий предок современных носителей гаплогруппы I2a на Балканах жил всего 2300 лет назад, в конце прошлой эры, и совсем недавно это же было подтверждено расчетами по снипам группой YFull. Эти расчеты показали, что четыре основных балканских субклада (S17250, Z17855, Y4460 и A2512), которые за последние две тысячи лет уже разошлись от Греции до Балтики, включая Россию, Украину и Белоруссию, образовались каждый примерно 2200 лет назад.
Вывод простой – попгенетика не является исторической наукой, это наука сугубо описательная, хотя иногда и бывает полезной, а вот ДНК-генеалогия является наукой исторической – и по методологии датировок, и по их интерпретациям.
Еще урок, который можно вынести из рассмотрения ископаемых ДНК и их субкладов по территориям и датировкам – это то, что по ним нельзя делать скоропалительных выводов и заключений. Полезно нанести соответствующие данные на карты, как это сделал И.Л. Рожанский, но связывать полученные точки прямыми линиями и утверждать (или даже предполагать), что именно так шли древние миграции, как это делают многие в сетевых «дискуссиях» – это, откровенно говоря, профанация. Нельзя по одной точке утверждать, что она адекватно представляет по составу соответствующую археологическую культуру. Это же нельзя делать и по двум-трем точкам, найденным в одном регионе, если эти две- три точки представляют разные гаплогруппы. Надо обратить особое внимание, найдены ли те два-три образца в одном захоронении или в разных, обратить внимание на археологические датировки каждого образца, они могут различаться на сотни, а то и на тысячи лет, обратить внимание на похоронный обряд, а именно на положение костяка, что является одним из самых устойчивых археологических признаков, и может свидетельствовать, одна ли это культура, или разные. Наконец, надо внимательно проверять источники возможных ошибок в определении гаплогрупп и субкладов, поскольку попгенетики славятся своей неряшливостью и использованием устаревшей номенклатуры, что может приводить к фактическим ошибкам в типировании, и, как результат, приводить к принципиально искаженным историческим интерпретациям.
Примеров всего этого – множество. Использование неверных «скоростей Животовского» привело коллектив под руководством Underhill (2010), в составе которого были и Животовский, и Балановские, и эстонские попгенетики, к выводу о том, что арии в Индию не переходили, поскольку по их «расчетам» это должно было быть не позже 10 тысяч лет назад, «а ариев тогда не было». Эти же «скорости Животовского» на протяжении ряда лет использовались как доказательство, что арии – это выдумка английских колонизаторов, для унижения индийского народа, и что гаплогруппа R1a образовалась в Индии 13-15 тысяч лет назад. Эти же «скорости Животовского» привели коллектив попгенетиков под руководством Tyler-Smith (2010) к тому, что разделение христиан и мусульман на Ближнем Востоке произошло якобы 3475 лет назад. Поскольку авторы (в числе которых был также известный Спенсер Уэллс) отмечают, что это намного раньше времен как христианства, так и мусульманства, то они делают вывод, что эти народы стали разделяться еще в те времена, то есть за полторы тысячи лет до христианства, не говоря о мусульманстве. Потрясающе! Они, так сказать, чувствовали, что через две тысячи лет им придется худо. На самом деле, разделив на 3, тот самый средний поправочный коэффициент, отличающий «коэффициент Животовского» от здравого смысла, они бы получили, что разделение христиан и мусульман произошло примерно 1100-1200 лет назад, то есть в 8-10 вв. нашей эры, именно во времена распространения мусульманства.
Еще один урок дает совсем недавняя публикация, по ископаемым ДНК на Пиренейском полуострове, рассмотренная в части 3 этой серии статей. Согласно публикации (Martiniano et al., 2017), в двух захоронениях среднего бронзового века (по датировкам авторов 3740-3430 лет назад) нашли три гаплогруппы R1b. К сожалению, снипы авторы не привели, а привели, делая типичную ошибку попгенетиков, только названия субкладов, которые меняются каждый год и чаще, и остается только гадать, какие снипы на самом деле у тех R1b1a2, R1b1a2a1a2 и R1b1a2a1a2, которые они показали. Если названия субкладов современные, 2017 года, то первый – это R1b-V88, а двух последних в классификации вообще нет, тем более что они должны быть нижестоящими от V88, номенклатура которых крайне ограничена. В общем, нет таких. Но загадка довольно быстро решилась, авторы действительно использовали устаревшую номенклатуру, которая была актуальна в 2013-2015 годах, в 2016-м она уже поменялась, и опять поменялась в 2017-м. Короче, один из R1b оказался M269, и два других P312. Всё стало на свои места.
Действительно, как было описано еще в статье в Advances in Anthropology (Klyosov, 2012), носители R1b-M269-L11 (он же L151 и он же P311) прибыли примерно 4800 лет назад на Пиренеи, и от них образовались P312 и U106. Датировка общего предка для современных 829 гаплотипов R1b-U106 в 111-маркерном формате равна 3584±359 лет назад (без округления, Klyosov & Kilin, Adv. Anthropol., 2016), такая же датировка относится и к общему предку современных R1b-P312, как было показано еще в 2009 году (Klyosov, J. Gen. Geneal.) на 464 гаплотипах, и с тех пор многократно воспроизводилась. И вот теперь те же R1b-P312 найдены в захоронении в Португалии с археологической датировкой 3740-3430 лет назад, что практически совпадает с расчетами ДНК-генеалогии. Там, правда, в расчетах есть особенности, что будет обсуждаться ниже.
Вывод – всё тот же, нельзя скоропалительно «опровергать» то, что было доказано вдоль и поперек раньше, тем более на основании единичной точки, которая к тому же оказывается ошибочной. И вот к чему это приводит – в статье И.Л. Рожанского по гаплокартам древности (4500-4000 лет назад) сообщается – «Из 9 образцов, в том числе 5 из Страны Басков, оказался только один носитель гаплогруппы R1b… без подтвержденного снипа М269. Впрочем, его данные еще не прошли проверку у экспертов, а потому следует дождаться результатов их анализа. Остальные образцы, как и на предыдущей карте, представляют гаплогруппы «Старой Европы», хотя по археологическому контексту они из культуры колоколовидных кубков». Судя по тексту рядом, речь идет о статье Olalde и др. (2017) о культуре колоколовидных кубков, но тогда данные Martiniano и др., о которых речь выше, просто не учтены, а они добавляют три образца субклада R1b-M269 – один сам M269, и два M269-P312. Как видим, интерпретация по сравнению с той, которую (правда, в уклончивом виде) дал И.Л. Рожанский, резко меняется. И все из-за того, что или не учтены новые данные (Martiniano, 2017), или из-за ошибки в типировании образцов, которые оказались R1b-M269 и (дважды) R1b-P312. Надо сказать, что И.Л. Рожанский не дал категорических интерпретаций по Пиренеям, но его уклончивые комментарии, что R1b там практически (или почти) нет, а в культуру колоколовидных кубков попали «по археологическому контексту» гаплогруппы «Старой Европы», видимо, G2, I и I2a1, уже разбежались по другим комментаторам, значительно более категоричным, которых хлебом не корми, а только дай «разрушить» то, что годами создавалось, и никакого «разрушения» там нет и быть не может.
Что же там за «археологический контекст», о котором вскользь, без пояснений, говорит И.Л. Рожанский? Авторы (Olalde и др., 2017) не дают никаких критериев, на основании которого они отнесли 54 ископаемых образца к «ассоциированным с культурой колоколовидных кубков (ККК)», хотя повторили этот термин «ассоциированные с ККК» в статье 24 раза. Среди них на Пиренеях были один G2 (с датировкой 3915±29 лет назад), четыре I2a2 (два из них с датировками 4051±28 и 3775±25 лет назад, остальные датировки не сообщались), два R1b1 (один, R1b1-L278, с датировкой 3965±29 лет назад). То, что носители гаплогрупп G2 и I2a были в Европе с древних времен, хорошо известно, как и известны их ранние находки с датировками 5000 и 7000 лет назад, еще до прибытия эрбинов на Пиренеи. Поскольку критерии, на каком основании носители найденных G2 и I2a, были зачислены в ККК, неизвестны, тем более когда основной состав ККК давно, многие века до того ушел на север, в континентальную Европу, то оставим это зачисление на совести авторов статьи. Но что примечательно, авторы статьи отмечают, что насельники ККК в континентальной Европе на 84% имели гаплогруппу R1b, и у тех ископаемых образцов, у кого определили более глубокий субклад, он оказался R1b-P312. Так что с отнесением ККК к гаплогруппе R1b, во всяком случае преимущественном, все в порядке, как и с тем, что ККК вышли с Пиреней с преимущественным субкладом P312.
Еще урок – что отсутствие археологических данных вовсе не означает, что миграций в тех краях не было. Наглядный пример – то, что гаплогруппа R1a-Z645-Z93-Z2123 найдена в цепи археологических культур на восток по Евразии, до Уральских гор и далее до Алтая, и она же найдена в обилии в Индии, включая высшие касты Индии, показывает, что переход ариев, носителей этой гаплогруппы, в Индию, имел место. Но никаких археологических данных на этот счет найдено пока не было. Это дало повод некоторым (маргинальным, как правило) историкам провозгласить, что арии в Индию не переходили, и даже придумать словечко «инвазионист» для тех, кто со всей очевидностью считает, особенно основываясь на данных ДНК-генеалогии, что такой переход был.
Здесь же – что датировки археологов в отношении археологических культур далеко не означают, что те культуры правильно датированы. Пример – срубную культуру, она же срубная культурно-историческая общность, археологи датируют временами 3600-3200 лет назад. Костные останки из срубной культуры в Самарской области, датированные 3550±300 лет назад, и были выбраны для палеогенетических исследований, которые показали гаплогруппы R1a-Z645-Z93 и R1a-Z645-Z93-Z2123, то есть те же самые, что найдены в Индии, в том числе в высших кастах. Но данные ДНК-генеалогии показывают, что движение на восток ариев, с гаплогруппой R1a-Z645 и нижестоящих субкладов, началось примерно 4500 лет назад, и действительно, синташтинские образцы (Южный Урал) показали ту же гаплогруппу, с датировками 4060-4313 и 3911-4141 лет назад. Это означает, что либо датировки срубной культуры занижены археологами на 500-800 лет, и должны быть в диапазоне 4400-4200 лет назад, либо соответствующая культура ариев в их продвижении на восток просто упущена, не найдена. Действительно, датировка образования снипа Z2123 – 4200±400 лет назад (https://www.yfull.com/tree/R1a/). То, что найдено и названо «срубной культурно-исторической общностью» – это относительно недавний «хвост» арийских миграций. То же самое и с хвалынской культурой, она должна датироваться не позднее 8 тысяч лет назад, и с ямной, там датировки должны быть не позднее 6-7 тысяч лет назад. Субклад R1b-Z2103, найденный в захоронениях ямной культуры, образовался 6200±800 лет назад (https://www.yfull.com/tree/R1b/). Те же 6-7 тысяч лет назад носители гаплогруппы R1b были уже на Кавказе, в ходе своего продвижения в Месопотамию.
Еще один урок, сопряженный с предыдущим, который дает рассмотрение ископаемых образцов ДНК – то, что если их пока не нашли, то это не значит, что их не может быть вообще. Отсутствие (пока) доказательств не есть доказательство отсутствия, об этом знает любой криминалист. Об этом много говорилось в части 3 этой серии статей. Но приходится повторять, потому что даже после прочтения той статьи, в ходе сетевых обсуждений, комментаторы опять сваливаются в ту же порочную колею, несмотря на сделанные предостережения. В ответ на мои слова в дискуссии, что «Ничего описанные палеогенетиками новые образцы и определенные гаплогруппы не «разрушают», они или дополняют ранние схемы, или их подтверждают», и о том, что «При малом числе определений нельзя считать, что пока не найденных гаплогрупп там нет вообще», некоторые продолжают педалировать негативности в обсуждении. Вот – прямая цитата с форума «ДНК-генеалогия как историческая наука» – «А все-таки на Балканах по-прежнему не найдены ископаемые R1a, все древние R1a Европы найдены значительно северней Балкан. Если так и не найдут, то, на мой взгляд, это опровергнет гипотезу, что арии разошлись по Европе с Балкан». Другой подхватывает – «более того, возможен пересмотр гипотезы о передвижении носителей R1a по южной дуге, через Иран и Анатолию. Более вероятно перемещение их с востока в Европу по… пути севернее Арала, Каспия и Понта». Откуда это «более вероятно» взяли, при полном отсутствии данных – остается только удивляться. На торопливых воду возят.
Прекрасно понимаю, что все подобные скоропалительные «соображения» можно, и даже нужно игнорировать, но беспокоит то, что они звучат на ресурсе под названием «ДНК-генеалогия как историческая наука». Беспокоит то, что участники столь полны рвения «опровергать» и «пересматривать», не имея на этот счет ровно никаких оснований. Это – плохой знак еще и потому, что эти торопливые, похоже, пренебрегают важным принципом под названием «приоритет первой гипотезы». Она является серьезной этической и конструктивной базой в науке. Кто-то проработал разные варианты интерпретаций, и выбрал наименее противоречивую. Следует исходить из того, что при этом была проведена большая работа, зачем от нее автоматически отказываться? Эта гипотеза должна быть при последующем рассмотрении первой (она может быть улучшенной редакцией предыдущей, но это в данном контексте не имеет значения). Данная концепция (гипотеза) должна быть приоритетной для рассмотрения. Если она не выдерживает проверки другими данными (старыми, известными, или новыми), то вырабатывается новая гипотеза, при максимальном сохранении старой, если это возможно. Если же «первая гипотеза» выдерживает проверку новыми данными, или новые данные ее дополняют, то «первая гипотеза» дополняется, модифицируется. Она становится лучше, более продвинутой. Это – мой путь, которого я стараюсь придерживаться.
Перейдем к гаплогруппе R1a, а именно к «приоритету первой гипотезы». Она много раз публиковалась и обсуждалась, поэтому буду относительно краток. Гаплогруппа R1a образовалась 22±2 тысяч лет назад, предположительно в Южной Сибири, в Алтайском регионе, и по «южной дуге» прошла через северо-западный Китай, Тибет, Иран, Анатолию на Балканы, расселилась в Европе, и примерно 5000-4500 тысяч лет назад продвинулась из Европы на Русскую равнину. Это – датировка как R1a-Z280, так и R1a-Z93. Дальнейшие пути миграций известны.
Вспомогательные свидетельства к тому – архаичные гаплотипы в Тибете, Гималаях, в северо-западном Китае у уйгуров, на Ближнем Востоке, на Коморских островах у африканского побережья (там тоже архаичные гаплотипы R1a, с общим предком ранее 8 тысяч лет назад), обнаружение прото-ИЕ языка в Анатолии с датировкой 8700±800 лет назад (эта датировка дается в известной статье Gray and Atkinson (Nature, 2003), и с ней в целом совпадают другие датировки многих лингвистов), и на Балканах («балканская теория происхождения ИЕ языка»), а также в южных степях Русской равнины с намного более поздней датировкой, которая, впрочем, никогда не была четко изложена, даже с погрешностью. Еще данные к тому – обнаружение в Европе архаичных образцов R1a, гаплотипы которых имеют архаичные же DYS392 = 13, в то время как почти все остальные имеют там «11». В Европе их найдено уже 50 образцов (древнейших субкладов YP4141 и YP1272, образовались 18300 и 14100 лет назад, соответственно), все у наших современников (включая из России и Белоруссии), а также в Иране (5 образцов), в Турции (3 образца), Катаре и Йемене. Заметим, что Иран, Ближний Восток и Аравийский полуостров не противоречат «южной дуге» миграции R1a, как и R1a на Коморских островах. Не противоречит этому и недавняя находка современного носителя архаичной R1a-YP1272 в Египте (https://www.yfull.com/tree/R-YP1272), а также нижестоящего субклада R1a-YP1272-YP1276 в Тунисе, такого же нашли в современной Белоруссии.
Итак, это «приоритетная первая гипотеза» миграций R1a, и любой последующий анализ и выдвигаемые гипотезы, в том числе с использованием ископаемых ДНК, обязаны проводить с ней состыковку, и либо отдельные положения опровергать, если ранг новых свидетельств выше, или эту гипотезу дополнять. То, что ископаемых ДНК где-то не нашли, не является свидетельством более высокого ранга, и не может быть основанием для ревизии первой приоритетной гипотезы. Всегда необходимо пытаться встраивать новые данные в первую приоритетную гипотезу, хотя бы потому, что она первая, и игнорировать этот факт неэтично. Например, обнаружение ископаемых гаплогрупп в Прибалтике (в культуре ямочно-гребенчатой керамики в Эстонии) и в Карелии следует интерпретировать так, что они прошли с Балкан на север, в Прибалтику. Разумеется, если будут найдены конкретные свидетельства, что гаплогруппа R1a прошла, например, из Сибири прямиком в Прибалтику, то первая приоритетная гипотеза будет модифицирована соответствующим образом. Балканский путь при этом останется до получения свидетельств, что носители гаплогруппы R1a туда не приходили, что ИЕ языки через Анатолию на запад не проходили, что балканская теория ИЕ языков неверна, причем не на уровне мнений или альтернативных вариантов, а на уровне прямых доказательств.
Перейдем к гаплогруппе R1b, опять в качестве «приоритета первой гипотезы». Эта гипотеза была высказана около десяти лет назад, и с тех пор никаких противоречий с новыми данными в ней не обнаружено. Дополнения, естественно, появлялись по мере идентификации новых и новых субкладов. Никто, понятно, не мог предположить, что в ямной культуре, помимо гаплогруппы R1b, предложенной ранее (Клёсов, 2009, 2010, 2012), будет выявлен субклад L23, и далее нижеследующие субклады L23 > Z2103 > Z2106 > Z2108 > KMS67 (последние три образовались примерно 6000 лет назад, то есть именно во времена ямной культуры), которых в Европе практически нет, и это вбило окончательный гвоздь в гроб старых представлений М. Гимбутас о прямом переходе степняков из ямной культуры в Европу, да еще к тому же якобы с индоевропейским языком.
По той первой гипотезе эрбины, носители гаплогруппы R1b, продвигались в период (предположительно) 12-8 тысяч лет назад из Южной Сибири по «северной дуге», через Южный Урал и Северный Казахстан, создав культуры Тургайского прогиба, суртандинскую, ботайскую, хвалынскую, ямную, оттуда – через Кавказ в Месопотамию. На всем этом пути встречается положение древних костяков на спине, характерное для носителей R1b, но не R1a. Археологи дают хвалынской культуре времена 7000-5500 лет назад (на самом деле должно быть древнее), ямной культуре 5600-4300 лет назад (датировки тоже явно заниженные ), и после заселения (части) Русской равнины эрбины направились на Кавказ (не позднее 7000 лет наад) и далее в Месопотамию и на Ближний Восток, основали государство шумеров (6500-6000 лет назад), и после того направились на запад, через Египет, вдоль южного берега Средиземного моря до Атлантики, переправились через Гибралтар на Пиренейский полуостров около 5000 лет назад, основали там культуру колоколовидных кубков (примерно 4800 лет назад), далее направились на север, в континентальную Европу, и заселили ее в течение последующих 1000-1500 лет, между 4500 и 3000 лет назад. В ходе этого заселения из Европы пропали почти все мужские автохтонные гаплогруппы, включая С, E1b, F, G2a, H2, I1, I2a, I2c, J2, K, R1a, и только R1b интенсивно приумножалась, как приумножались и автохтонные женские мтДНК. С тех пор гаплогруппа R1b продолжает оставаться доминирующей по численности гаплогруппой центральной и западной Европы, которая в настоящее время там достигает 60%. На 95% (или более) это потомки двух пиренейских субкладов R1b-P312 и R1b-U152.
Люди, игнорирующие приоритет первой гипотезы, что уже показывает их неквалифицированность, приводят как «аргумент» то, что ископаемых гаплогрупп R1a на Балканах пока не нашли. Хорошо, давайте посмотрим внимательно на этот «аргумент». Не будем повторять, что «пока не нашли» – это вообще не аргумент, ни в пользу того, что они были, ни в пользу того, что их не было. Это вообще не аргумент, это «текущий факт». А вот данные. Согласно статье Mathieson и др. (2017), в балканской культуре Лепенского Вира (в Сербии) обнаружено более 200 захоронений, из них палеогенетики работали только с двумя. В одном было пять скелетных останков, из них выбрали женский, с датировкой 7474±35 лет назад (в сводной таблице в Приложении к статье указано 8006 лет назад), Y-хромосомной гаплогруппы там, разумеется, нет. В другом – мужской скелет с датировкой 7670±35 лет назад (в Приложении указано 8017 лет назад), гаплогруппа R1b1a-L754, положение костяка – в вытянутом виде на спине. Это – характерное положение костяка для носителей гаплогруппы R1b, от мезолита (или раннего неолита) до сарматских времен, I тыс. н.э. Обнаруженный субклад – архаичный, он образовался примерно 18900 лет назад, и является вторым нисходящим от R1b в иерархии снипов R1b-M343 > L278 > L754.
Вот и всё по Лепенскому Виру. Из более чем двухсот захоронений. Согласитесь, что торопиться с выводами о том, какие гаплогруппы «там не нашли» – это несколько преждевременно. А вот и другие данные, которые в данной статье не упомянуты. В статье Boric & Dimitrijevic (2007) многократно (12 раз) упоминается о костяках Лепенского Вира в скорченном положении, совершенно не характерном для эрбинов, как и для носителей гаплогруппы I2, тоже обычно захороненных на спине (см. ниже). Можно вдаваться в детали, чем различаются захоронения на спине для R1b и I2, например, в положениях рук (вдоль тела или сложены), но с этим надо работать. Скорченное положение костяка на боку характерно для племен R1a, мужчины – на правом боку, женщины на левом, с противоположной ориентацией направления головы (Клёсов, 2015, 2016, ссылка). Именно эти данные о скорченном положении костяков в культуре Лепенского Вира явились основанием для первой гипотезы о прибытии носителей R1a на Балканы 8-10 тысяч лет назад, в совокупности со вспомогательными данными об обнаружении прото-ИЕ языка в Анатолии и на Балканах в те же времена.
Полагаю, торопливость «энтузиастов разрушения» очевидна – они выхватывают буквально одну «экспериментальную точку» из общей картины, которой не знают, да и знать не хотят, и пытаются делать (или хотя бы предполагают) глобальные выводы.
Ясно то, что эрбины в Лепенском Вире и в округе были. На расстоянии всего пяти километров от Лепенского Вира находится местечко Падина, тоже в Сербии. Там зафиксировано 33 захоронения, из них палеогенетики рассмотрели 12 образцов ископаемых ДНК, десять из которых описаны в Приложении к статье Mathieson и др. (2017), из них шесть женских и четыре мужских. Мужские – три гаплогруппы R1b, все – субклада R1b-L754, того же, что и в Лепенском Вире, и одна – I2a1-P37 (образовалась примерно 21200 лет назад). Датировки для R1b были 10835, 10425 и 9800 лет назад, для I2a1 – 10493 лет назад, то есть все на 2-3 тысячи лет древнее, чем в Лепенском Вире. Положение костяка было на спине для одного R1b (для двух других не установлено), и для единственного I2a1. Это – захоронения под номерами 9, 12, 22 и 14. Отмечено, что в захоронении 30 положение костяка было скорченным, но гаплогруппу его не определяли.
Всего в трех километрах от Лепенского Вира есть местечко Власак, в нем тестировали 17 захоронений, из них 9 женских скелетных останков и 8 мужских, семь гаплогруппы I и нижестоящих субкладов, и один – R1b1a-L754, как и в Лепенском Вире и Падине, с датировкой 8450 лет назад, примерно на сто лет древнее, чем в Лепенском Вире, но на 1500-2000 лет «моложе», чем в Падине. Среди группы I два образца были самóй исходной гаплогруппы I-M170, остальные – I2-M438, I2a2a-M223, I2a2a1b-CTS10100, и два I2a2a1b2-Z161. Интересно, что почти все костяки гаплогруппы I и нижестоящих субкладов, для которых положение удалось установить, были на спине, как и костяки гаплогруппы R1b. На спине были захоронены и все женские костяки в тех 17 захоронениях. Датировки костяков гаплогруппы I-M170 и I2 – самые древние в этих захоронениях, 10040, 8325 и 8390 лет назад, нижеследующих субкладов – 8325 и 8000 лет назад (последняя датировка – у четырех костяков).
Еще древние захоронения в Сербии включают три образца балканского неолита с гаплогруппой G2a (местечко Gomolava, датировки 7440, 6557 и 6483 лет назад) и четыре образца мезолита-неолита с гаплогруппой I2a (местечко Hajdučka Vodemica, датировки 8123 и три образца 8600 лет назад).
Интересным явилось обнаружение гаплогруппы Н2 на Балканах, куда они явно пришли из Анатолии. Из семи ископаемых образцов Н2, один был найден в Леванте с самой древней датировкой (8700 лет назад), три – в Анатолии (все 8300 лет назад), один в балканской культуре Старчево (7569 лет назад), один в Болгарии (5132 лет назад), и один в Иберии (5700 лет назад). Это интересно и потому, что в настоящее время гаплогруппа H встречается в основном в Индии, Пакистане, Шри-Ланке, но исключительно субклада H1, он же – основной у цыган Европы. Потомки субклада H2, найденного в захоронениях европейского мезолита и неолита, встречаются в настоящее время в Европе (Италия, Франция, Англия, Швеция), Турции и в странах Персидского залива (Кувейт, Катар). В сети есть масса фантазий про приход дравидов (иногда с указанием их гаплогруппы, якобы Н) в Европу, правда, во времена бронзового века, как якобы спасавшихся от прибывших в Индию ариев, но эти фантазии ничем не обоснованы. С гаплогруппой Н1 дравидов в Европе тоже не складывается, такую не нашли, во всяком случае, пока.
Теперь, когда общий обзор «приоритетных гипотез» завершен, как и типичных уроков при расмотрении ископаемых образцов ДНК, рассмотрим самые новые данные по ископаемым ДНК (по африканцам и минойцам), и затем приступим к обобщениям, а также к тому, какие изменения целесообразно внести в «приоритетные гипотезы».
В июне этого года опубликована статья группы авторов из Швеции и Южной Африки, под названием «Древние геномы из южной Африки помещают разнообразие современного человека глубже 260 тысяч лет назад». Датировка была получена сопоставлением числа мутаций у ископаемого образца ДНК из Южной Африки с археологической датировкой 1900 лет назад, и современных людей, с использованием константы скорости мутации 1.5х10-8 на нуклеотид на поколение в 30 лет, то есть 0.5х10-9 на нуклеотид в год. Вообще-то специальные исследования недавно пришли к более надежной константе 0.8х10-9 на нуклеотид в год, то есть у авторов статьи скорее всего завышение датировки в 1.6 раз, и должно быть примерно 160 тысяч лет назад, то есть то, что было вычислено и описано нами еще пять лет назад для расхождения ДНК-линий тех, кто сейчас живут в Африке, с неафриканцами (Klyosov & Rozhanskii, 2012, Advances in Anthropology), но попгенетики это, видимо, не читатели, они, как известно, писатели, а не читатели. Тем не менее, авторы недавней статьи так воодушевились этой полученной датировкой, что повторили ее в статье 10 раз. Но если говорить не про «разнообразие современного человека», а про образование наиболее древних ДНК-линий гаплогруппы А00, то там датировки варьируются от 208 до 338 тысяч лет назад (Elhaik и др., 2014; Mendez и др., 2014), 217 тысяч лет назад (Klyosov & Kilin, 2016), и 236 тысяч лет назад (https://www.yfull.com/tree/A00/). Так что ничего нового в этом отношении авторы статьи про древние геномы не сообщили, хотя это у них главный вывод.
Но нас сейчас интересуют не столько их геномные данные с неизменными продолжительными и вязкими рассуждениями про «геномную примесность» ископаемых африканцев 2000 и 400 лет назад, хотя это, похоже, вообще никого не интересует, тем более понимая, как это было получено, нас интересуют их гаплогруппы и субклады. В обсуждаемой статье были рассмотрены семь ископаемых образцов ДНК из Южной Африки, из них пять женских (у всех мтДНК были разными, но это дело обычное, хотя и относились к двум группам – L0d и L3e), и два мужских, гаплогруппы A1b1b2 в обоих случаях. Датировка обоих мужских костных останков была тоже практически одинаковой, и пересекалась в пределах погрешности, примерно 2000 лет назад. Женские костяки датировались в пределах 300-500 лет назад. Чтобы понять, где на дереве субкладов гаплогруппы А (там на самом деле серия гаплогрупп под общим индексом А), расположен этот ископаемый субклад A1b1b2, пометим его желтым цветом:

Этот субклад, A1b1b2-L427, образовался примерно 54900 лет назад (https://www.yfull.com/tree/A1b1/), и в статье, разумеется, недотипирован. Где он образовался – неизвестно, вполне может быть, что и за пределами Африки, никто этого в самом деле не знает. Датировок образования субклада в статье, конечно, тоже нет. Там главный вопрос – «примесность геномов», хотя так и не объяснено, кому и зачем это надо, и что это дает.
 «Кластерный анализ» («примесность») для четырех женщин – «фермеров железного века», по определению авторов рассматриваемой статьи (вертикальная панель слева), датировки 280-530 лет назад, и трех образцов – «охотников-собирателей» каменного века (вертикальная панель справа), из которых два мужских скелета (с гаплогруппой A1b1b2-L427, пол третьего не определен, датировка 1800-2300 лет назад. Величины индекса К (пояснение в тексте) на панелях сверху вниз равны соответственно 3, 5, 7, 12, 13.
«Кластерный анализ» («примесность») для четырех женщин – «фермеров железного века», по определению авторов рассматриваемой статьи (вертикальная панель слева), датировки 280-530 лет назад, и трех образцов – «охотников-собирателей» каменного века (вертикальная панель справа), из которых два мужских скелета (с гаплогруппой A1b1b2-L427, пол третьего не определен, датировка 1800-2300 лет назад. Величины индекса К (пояснение в тексте) на панелях сверху вниз равны соответственно 3, 5, 7, 12, 13.Пример изучения «примесности» приведен на диаграмме слева. Панели сверху вниз соответствуют величинам индекса «К», равному 3, 5, 7, 12 и 13. Эти индексы соответствуют «спектральному» разложению генома на число «компонентов», которые можно условно рассматривать как число предков данного генома. Видно, что для ископаемых ДНК женщин – «фермеров железного века» несколько веков назад картина «спектрального» разложения одинакова для числа предков от 3 до 7, но при 12 предках картина резко меняется и становится вообще однородной. При добавлении еще одного предка картина опять резко меняется, и состав предков уже опять совершенно другой. Темно-зеленый цвет приписывается авторами статьи современному племени йоруба, светло-зеленый, видимо, носителям языка банту, хотя там есть неясности, желтый – современным людям Karretjee из племени San из южной части Южной Африки, но проблема в том, что этот цвет появляется у них только при К=13. Опять неясно, зачем и кому это нужно. Похоже, что это нужно «широкогеномным» исключительно для описательности, так, зафиксировать и забыть, поскольку никаких выводов из этого не делается.
Из диаграммы видно, что никаких параллелей или другой связи между женщинами железного века (слева) и мужчинами каменного века (справа) не прослеживается. Красный цвет – это, похоже, современные люди из племени San из северной части Южной Африки, хотя и там проблема – уже при К=7 их цвет на диаграмме резко меняется от сплошного красного к сплошному синему (на этой диаграмме не показано, поскольку общая диаграмма содержит 312 панелей, и все столь же бессмысленны), а у ископаемых образцов каменного века (справа) только следы этого синего цвета. Для общей «информативности» сообщим, что при К=13 люди каменного века от почти абсолютного соответствия северным San (при К=3) переходят к почти абсолютному соответствию южным San. И вот подробными описаниями этой, мягко говоря, ерунды заполнена практически вся рассматриваемая статья.
Возвращаясь к субкладу A1b1b2-L427, отметим, что где бы он ни образовался десятки тысячелетий назад, все субклады, нижестоящие от A1b, включая L427, у неафриканцев отсутствуют, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т.
Так или иначе, тот факт, что носитель субклада A1b1b2-L427 обнаружен в Южной Африке с датировкой 2000 лет назад, ничего, к сожалению, не дает для понимания древних миграций. И уж тем более ничего не дают «данные по примесности», которые варьируются как угодно при изменении условий расчетов. Здесь мы приводим эти данные просто «для коллекции».
В начале августа этого года в журнале Nature вышла статья группы авторов из 32 человек, под руководством David Reich из Гарвардской медицинской школы, под названием «Генетическое происхождение минойцев и микенцев». Статья, как и все работы, вышедшие из лаборатории Reich, является типичным попгенетическим исследованием, со всеми принципиальными проблемами «широкогеномных» публикаций. Мы на них кратко остановимся ниже, а сейчас упомянем, кто такие минойцы и микенцы, и какие у них были найдены гаплогруппы. На мой взгляд, гаплогруппы, приведенные в небольшой таблице в приложении к статье, и есть самый главный результат работы коллектива авторов. Всё остальное – мутные, вязкие, неопределенные сентенции о возможных «примесностях» в геномах минойцев и микенцев. Примеры также приведены ниже.
Итак, минойцы – это представители эгейской цивилизации острова Крит, которая датируется временами 4700-3400 лет назад. Минойцы – это не самоназвание тех людей, термин «минойцы» был придуман в конце 19-го века британским археологом Артуром Эвансом. В связи с этим можно только с иронической усмешкой вспоминать тех непримиримых «борцов с древними ариями», которые напирают на то, что «ариями» их называть нельзя, потому что нет доказательств, что это было их самоназвание. На самом деле подавляющее большинство названий древних племен – не самоназвания, а названия, данные историками, от античных до современных. Можно подумать, что, например, скифы называли себя скифами, американские индейцы – американскими индейцами, китайцы – китайцами, минойцы минойцами, и так далее.
Микенцы – это представители той же эгейской цивилизации, ее крито-микенской культуры, с датировкой примерно 3600-3100 лет назад, то есть несколько более поздней по сравнению с минойцами. Микенскую цивилизацию еще называют Ахейской Грецией.
Теперь пора показать гаплогруппы минойцев и микенцев, опубликованные в рассматриваемой статье. Из 19 образцов ископаемых ДНК (минойские, микенские, анатолийские, греческий, критский) 14 – это женские мтДНК. У всех 19 образцов, мужских и женских, есть 14 вариантов мтДНК, все разные – иначе говоря, это разнообразие не слишком информативно. У 14 женских ископаемых ДНК есть 11 вариантов мтДНК, то есть вывод тот же.
Если обратиться к ископаемым Y-ДНК, то они следующие. Из трех минойских образцов один найден в префектуре Ираклион (Гераклион), гаплогруппа G2a2b2, датировка 4900-3900 лет назад. Два образца найдены в префектуре Ласити, гаплогруппы J2a1 и J2a1d, датировки обоих образцов 4000-3700 лет назад. Положение Крита приведено на карте ниже (с его четырьмя префектурами), там же показан Ираклион, префектура Ласити примыкает к нему с востока.
Микенский – один ископаемый образец, из Греческого Пелопоннеса, гаплогруппа J2a1, как и у двух минойских, датировка 3700-3200 лет назад, то есть несколько более недавняя по сравнению с минойскими. Еще один образец – анатолийский (нынешняя Турция), гаплогруппа J1a, датировка 3925±35 лет назад.
Вот, собственно, и всё, что есть конкретного в обсуждаемой статье. Остальное – расплывчатые рассуждения. Если на время забыть о «широком геноме», то на что указывают гаплогруппы G2a и J2a у минойцев и микенцев с Крита и континентальной Греции? Да на древнюю Европу, Средиземноморье и Анатолию, ближайшее окружение Греции и Крита, а также на Иран и восточнее. Мы уже знаем, что гаплогруппа G2a была одной из основных в мезолите и палеолите Европы, и сейчас ее доля на Крите 10%, в Греции 7%, на Эгейских островах 9%, в Турции 11%, в Армении 12%, в других странах Кавказа до 75%. Ее немало по всей дуге от Гиндукуша до Анатолии – у калашей в Южном Гиндукуше ее 20%, у пуштунов Афганистана 10%, в Иране 10%. Гаплогруппы J2a еще больше по той же дуге – в Иране 23%, в Турции 24%, в Армении 22%, в Греции 23%, на Эгейских островах в среднем 20%, на Крите 34%. Еще больше ее на Ближнем Востоке, среди арабов и евреев. Теперь посмотрим, что на этот счет говорят «широкогеномные» попгенетики.

Самые главные положения статьи обычно выносят в Абстракт. Что же вынесено туда в обсуждаемой статье? Приводим почти весь текст Абстракта:
Мы показали, что минойцы и микенцы генетически сходны, имея по меньшей мере три четверти их предковости от первых неолитических фермеров Западной Анатолии и жителей островов Эгейского моря. Остальная часть – в основном от древних популяций, имеющих отношение к Кавказу и Ирану. Однако минойцы отличаются от микенцев в том, что у первых есть дополнительная предковость от источника, имеющего отношение к охотникам-собирателям Восточной Европы и Сибири, привнесенных через источник, имеющий отношение к населению или евразийской степи, или Армении. Современные греки сходны с микенцами, но с некоторым дополнительным разбавлением ранней неолитической предковости. Наши результаты поддерживают идею непрерывности, но не изоляции в истории популяции эгейцев, до и после времен самых ранних цивилизаций.
Кто что понял? Очень информативно, не так ли?
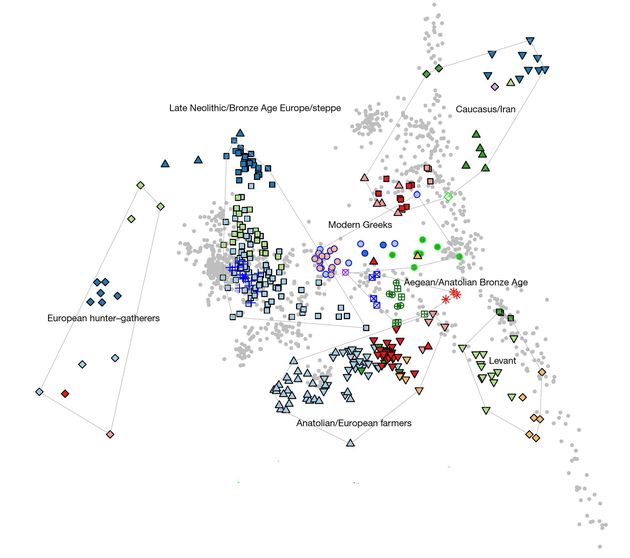
Добавим к этому «основания» для приведенных выводов в виде «диаграммы принципиальных компонент», приведенной выше. Обозначений координат, то есть что отложено по вертикали и что по горизонтали, здесь нет, потому что их нет вообще. Это – некие безымянные преобразования, которые рассчитал компьютер по сотням тысяч и миллионам снипов, и которые растянуты по вертикали и горизонтали. Их условно назвали «принципиальные компоненты». Каждая цветная точка – древний ископаемый образец ДНК, неважно, мужской или женский, и неважно, какой гаплогруппы, Y-хромосомной или митохондриальной. Каждая серая точка – современный геном. Всё рассмотрение и понимание (если таковое есть) основано на принципе «похожести», ближе точки – более похожи, то есть «родственные», у них «общие предки ближе», дальше – менее похожи. Но при желании и там можно «рассчитать» то, что общие предки «делают вклад». Минойцы и микенцы – близко к середине диаграммы, их символы – кружочки и квадратики с перекрестиями. Прямой крестик – минойцы (зеленые символы), косой крестик – микенцы (синие символы), они все окружены тонкой чертой с надписью «эгейцы/анатолийский бронзовый век», это потому что геномы анатолийцев бронзового века (красные звездочки) сидят на правом краю этой обводной линии, правда, не слишком близко. Чтобы их захватить, обводную линию удлинили в правую сторону.
Как видим, иранские образцы (справа вверху) сидят на максимальном удалении от минойцев-микенцев, как и «европейские охотники-собиратели» (слева). Треугольнички вершиной вниз – иранцы неолита, вершиной вверх – иранцы халколита (медного века). «Кавказские охотники-собиратели» – верхние дальние точки слева по обводной линии. Но эта удаленность не помешала авторам статьи отметить, что минойцы-микенцы имеют происхождение на четверть «от древних популяций, имеющих отношение к Кавказу и Ирану». Так, видимо, решил компьютер, хотя любой попгенетик знает (или должен знать), что сходство в фрагментах геномов, которые компьютер ищет и находит, имеет по меньшей мере три объяснения – сходство по общему происхождению, сходство просто по случайной похожести, и сходство по причинам одинаковой функции в геноме, поскольку все люди имеют одинаковые функции метаболизма и жизнедеятельности. Там сходство в геномных фрагментах просто обязано быть.
Поэтому параметры «сходства», выдаваемые компьютером, обычно весьма шатки, и их можно интерпретировать так или иначе. Это и есть то множество степеней свободы, которое было упомянуто выше. «Широкогеномным» попгенетикам остается только выбирать по вкусу вариант «происхождения». Отсюда и получается – «минойцы отличаются от микенцев в том, что у первых есть дополнительная предковость от источника, имеющего отношение к охотникам-собирателям Восточной Европы и Сибири, привнесенных через источник, имеющий отношение к населению или евразийской степи, или Армении», и прочее в их выводах, которые и составили Абстракт статьи. Включая заключительный перл – «Наши результаты поддерживают идею непрерывности, но не изоляции в истории популяции эгейцев, до и после времен самых ранних цивилизаций». Интересно, откуда появилась идея «изоляции»? Где такие вообще нашли?
Итог простой – никакой конструктивной информации данные обсуждаемой статьи по минойцам и микенцам не дают. Их гаплотипы, даже несколько, тут же прояснили бы картину. Но это не тот путь, по которому идут «широкогеномные» попгенетики.
в свете «приоритетных гипотез», выдвинутых ранее
Как сообщалось в первых трех частях данной серии статей, несмотря на обилие новых данных по ископаемым ДНК, они не привнесли значительных изменений в «приоритетные гипотезы» древних миграций, основанных на современной картине гаплогрупп, субкладов и гаплотипов по планете. Но они привнесли важные дополнения к нашим знаниям, основанным на «приоритетных гипотезах». Я называю их приоритетными, потому что они были высказаны первыми, и этические правила в науке требуют их рассматривать в первую очередь, прежде чем их опровергать или вносить измерения. Дополнять – это пожалуйста, и то при наличии новых данных, а не «мнений», «соображений», «логики» и прочих второстепенных и третьестепенных факторов. К новым данным, в том числе по ископаемым ДНК, тоже надо относиться с осторожностью, как было показано на ряде примеров в этой серии статей. Нередко бывает, что эти данные одиночные, ошибочные, приводятся с использованием устаревшей номенклатуры, и вообще некритически интерпретируются, поспешно, торопливо, с «налету».
Вот – очередной характерный пример. Приоритетная гипотеза говорит о том, что эрбины, носители гаплогруппы R1b, продвигались со стороны Южной Сибири на запад. Когда это движение (миграция) началось – мы не знаем, но мы знаем, что гаплогруппа R1b образовалась примерно 21 тысячу лет назад. В таблице YFull говорится, что 22800 лет назад, но эта датировка появилась как искусственное приравнивание к датировке образования гаплогруппы R1a, постулируя, что R1a и R1b образовались в одно и то же время. Это, конечно, несерьезный постулат, потому что образование R1a и R1b – совершенно независимые события. И, действительно, к гаплогруппе R1a по данным YFull ведут 169 снип-мутаций, а к гаплогруппе R1b – 145 снип-мутаций, что транслируется, соответственно, в 24400 и 21000 лет. В контексте настоящего изложения это не так важно, но торопливые любители уже строят «теории», исходя из того, что раз эти гаплогруппы образовались якобы в одно время, и, следовательно, якобы в одном месте, то и язык у них должен был быть якобы одинаковый. Но это непроверяемо, неверифицируемо, а значит, ненаучно. Тем более что они образовались, вполне возможно, с разницей в тысячелетия, и, вполне возможно, в разных местах.
Итак, движение гаплогруппы R1b на запад происходило когда-то от времени образования гаплогруппы или тысячелетиями позже, через Южный Урал, Северный Казахстан, возможно, через маханджарскую культуру примерно 10-9 тысяч лет назад (хотя данных о том, что это были именно эрбины, нет, и костных останков людей там пока не найдено; то, что там были эрбины, предположили археологи, копающие маханджарскую культуру), и через хвалынскую культуру до ямной, в период 8-7 тысяч лет назад. Эти датировки должны, видимо, согласоваться с датировками нарвской культуры на территории современной Латвии (8-5 тысяч лет назад), где были найдены захоронения эрбинов с датировкой 7690±100 и 7030±225 лет назад. Некоторые археологи утверждают, что из приволжских культур хвалынской и/или ямной, их насельники поднялись речными путями до Прибалтики (см. предыдущие части данной статьи).
Так вот, приоритетная гипотеза утверждает (или предполагает), что хвалынская и ямная культуры маркируют миграционный путь носителей гаплогруппы R1b от Южной Сибири до Волжских степей. Но вот опубликованы данные по трем образцам ДНК хвалынской культуры, и любители, как водится, дружно заголосили – ура, культура смешанная, там оказались три гаплогруппы – R1b, R1a, и Q1a, все в интервале датировок 7215-6015 лет назад, то есть в интеврале 1200 лет, дальше, чем от нас до Киевской Руси. Мне, признаться, никогда не было понятно, откуда у дилетантов такая страсть к опровержениям (как им кажется) гипотез и концепций, причем на основании не их, не своих данных, а чьих-то, опубликованных в других статьях. При этом в тех, новых данных, они особенно и не разбираются, их не анализируют. Главное, что – ура! – что-то разрушается! (как им кажется).
Они не задумываются, что носитель гаплогруппы R1a-М459 в хвалынской культуре захоронен не в скорченном положении на правом боку, как принято в культурах R1a, а на спине, в вытянутом положении, как принято в культурах R1b. Другими словами, соплеменники-хвалынцы рассматривали его не как «визитера» из чужой культуры, а как своего. А уж как он в хвалынскую культуру попал – мы не знаем и никогда, видимо, не узнаем. Может, «сын полка», может, пленный, который себя достойно проявил, может, младенцем нашли и воспитали – откуда нам знать?
Далее, дилетанты не задумываются, что следующая культура по пути миграции эрбинов, ямная, уже представила 11 ископаемых образцов, и ни один из них не показал гаплогруппу R1a или Q1a. Куда же они делись по ходу миграции? Не исключено, что они проявятся при последующих исследованиях палеогенетиков, но пока их нет. Из 11 образцов десять – R1b, и один – опять сторонний, но уже I2a, в Калмыкии, в отдельном захоронении, с датировкой, на 500-800 лет более поздней, чем R1b (для I2a – 4864-4158 лет назад). Так что встречаются ли сторонние гаплогруппы в ямной культуре? Да, встречаются, но, бывает, что в другое время и в другом месте, возможно, иногда вообще не относятся к ямной культуре. Поэтому торопиться и делать глобальные выводы вряд ли стоит. Во всяком случае, приоритетная гипотеза остается прежней.
Дадим относительно краткое изложение дополненной и расширенной приоритетной гипотезы древних миграций. Начнем с того, что где именно возник Homo sapiens и где и когда образовался анатомически современный человек – науке неизвестно. Некоторые антропологи считают эти два понятия эквивалентными, некоторые их разделяют. Если считать датировку образования древнейшей гаплогруппы A00 (примерно 240 тысяч лет назад) датировкой возникновения Homo sapiens, что вовсе не обязательно, то получим отправную точку современного человечества, но вряд ли это время появления анатомически современного человека. Скелетные останки предполагаемого Homo sapiens находят, хотя и крайне редко, с датировками 190-200 тысяч лет назад, но, как правило, они показывают архаичные особенности строения, что, по мнению многих антропологов, выводит их из вида Homo sapiens, и уж во всяком случае из категории «анатомически современного человека». По мнению ряда антропологов, анатомически современный человек появился не ранее 36 тысяч лет назад. Другие это оспаривают, и поднимают планку до 160 тысяч лет назад, или даже выше.
Никаких определенных данных о том, что Homo sapiens, как и анатомически современный человек, появились в Африке, нет. Неандерталец, «брат» Homo sapiens, определенно появился не в Африке, и следов его в Африке не обнаружено, хотя ищут более ста лет. Все «теории» о «выходе современного человека из Африки» 50, 60 или 70 тысяч лет назад, остаются фантазиями, это никогда не было показано, и все эти датировки никогда не были рассчитаны, они или придуманы, или натянуты с помощью совершенно необоснованных допущений (как описано в статье Klyosov & Rozhanskii, 2012, Advances in Anthropology). Выходит всё больше академических статей, в которых эти датировки либо отвергаются, либо просто не упоминаются, принимаясь по умолчанию неверными. Недавний пример – статья китайских авторов (2016) о нахождении в Китае костных останков древнего Homo sapiens c датировками 80-120 тысяч лет назад. В статье ни словом не упомянута Африка, но пересказ статьи в журнале Nature сообщает, что люди якобы вышли из Африки и мигрировали в Юго-Восточную Азию, где их останки и были недавно обнаружены с датировкой 80 тысяч лет назад. Про 120 тысяч лет назад пересказ статьи даже не упоминает. Это – типичный пример «обороны» представлений о «выходе современного человека из Африки», когда эти представления не подтверждаются прямыми данными.
Как бы там ни было, расчеты показывают, что современные африканские и современные неафриканские линии Y-хромосомы разошлись 160 тысяч лет назад (Klyosov & Rozhanskii, 2012). Здесь «африканские» линии называются не по месту их образования, а по месту современного проживания основного количества их носителей. На самом деле в Африке живут десятки миллионов носителей «неафриканских» Y-хромосом (в Южной Африке, в Египте, в Камеруне и Чаде, где живут миллионы носителей гаплогруппы R1b, и так далее), как и многие носители гаплогрупп серии А и В живут за пределами африканского континента. Более того, гаплогруппа В вовсе не образовалась из гаплогруппы А (любой из них), она образовалась из гаплогруппы ВТ, определенно за пределами Африки (Klyosov & Rozhanskii, 2012).
В период между 60 и 70 тысяч лет назад на Земле произошел катаклизм планетарного масштаба (Юрковец, 2016), огромная волна, стартовав в Тихом океане, обошла Землю, перехлестнула Антарктический материк, и ослабла при приближении к Балканам. Этот катаклизм привел к гибели большей части человечества, и в настоящее время гаплотипы всех мужских гаплогрупп сходятся к общему предку человечества, гаплогруппы ВТ, который жил 64±6 тысяч лет назад. Это согласуется с временами катаклизма, описанного выше. Исходя из описаний выше, а также из того, что наиболее древние скелеты анатомически современного человека найдены в Европе с датировками 45 тысяч лет назад (Англия, Италия) и 40 тысяч лет назад (Россия, Костенки), причем последний имел гаплогруппу С, можно предположить, что гаплогруппа ВТ пережила катаклизм именно в Европе, возможно, на Балканах. Ископаемые носители гаплогруппы ВТ найдены в Польше, гаплогруппы СТ- в Польше, Германии, Болгарии, Иордании, гаплогруппы С – в России, Испании, Болгарии, Хорватии, Бельгии, Австрии,Чехии, Румынии, Венгрии, Испании, Турции. Характерно, что гаплогруппа Н2, найденная в ископаемых образцах в Европе (Болгария, Венгрия, Испания), Турции и Израиле, не встречается в Индии, Пакистане и сопряженных регионах. Там наблюдается гаплогруппа Н1, которая также обычна среди цыган Европы. Это означает, что гаплогруппа Н2 не прибыла в Европу из Индии, а возникла в Европе (или поблизости) из гаплогруппы F, параллельной гаплогруппе С по филогении. Ископаемая гаплогруппа F была найдена в Испании.
То, что эти гаплогруппы действительно древнеевропейского происхождения, и их родоначальник пережил планетарный катаклизм 64±6 тысяч лет назад, могут показывать датировки их образования: гаплогруппа CF – 68500 лет назад, гаплогруппа С- 65900 лет назад, гаплогруппа DE – 68500 лет назад, гаплогруппа F – 65900 лет назад, гаплогруппа H – 48500 лет назад, и так далее (https://www.yfull.com/tree/BT/). Движение носителей гаплогрупп было, в частности, на восток, в центральную Азию, в Индостан, Юго-Восточную Азию, Полинезию, Австралию, намного позже в Америку, из центральной Азии. Ископаемых образцов ДНК там немного, но они информативны. Так, носитель гаплогруппы К (образовалась примерно 47200 лет назад, https://www.yfull.com/tree/BT/), уже 46900-43200 лет назад жил на берегу Иртыша в теперешней Омской области. Гаплогруппа Н (Н1) мигрировала в Индостан, ископаемые образцы ДНК там пока не получали, а гаплогруппа R оказалась в Южной Сибири, на берегу Байкала с археологической датировкой 24 тысячи лет назад (образовалась 31900 лет назад, https://www.yfull.com/tree/R/). По всей видимости, гаплогруппа Р оказалась там же, в Южной Сибири, так как из нее путем соответствующей мутации примерно 31900 лет назад образовалась гаплогруппа Q, носители которой ушли через сухую Берингию в Америку, прибыв туда по разным данным между 20 и 13 тысяч лет назад (Klyosov, 2009). В настоящее время до 95% индейцев Южной Америки (мужчин, разумеется) имеют гаплогруппу Q.
Гаплогруппа R по линии R1 образовала гаплогруппы R1a и R1b (24 и 21 тысяч лет назад, соответственно), которые разными путями в своей части мигрировали в Европу, как описано выше. Поскольку на своем пути по Евразии носители R1a и R1b мигрировали раздельно, если не считать случайных пересечений, как описано выше, их языки были разными, во всяком случае, между 9 и 3 тысячами лет назад. Первая датировка – это обнаружение прото-ИЕ языка в Анатолии, и далее на Балканах, определенно носителей гаплогруппы R1a, которые через тысячелетия принесли индоевропейские языки в Индостан, в Месопотамию (хетты) и на Ближний Восток (Митанни, Сирия), и оставив эти языки на Русской равнине, где современный русский язык имеет сейчас 54% совпадений базовой лексики (по стословнику Сводеша) с древнеиндийским языком (С.А. Старостин). Литовский язык имеет с древнеиндийским 53% совпадений, немецкий 57% (Старостин), но это все одно и то же в пределах погрешности расчетов.
Какой язык был у носителей R1a и R1b до 9 тысяч лет назад, мы не знаем, но нет совершенно никаких оснований считать, что эрбины пронесли через Евразию ИЕ языки, потому что и после этого они не оставляли ИЕ языков ни на Кавказе, ни на территории современной Турции, ни в Египте, ни у басков, ни на Британских островах до прибытия туда кельтских языков. Есть основания полагать, что язык носителей гаплогрупп Р > Q, R > R1 > R1b был предковым к тем, что недавно получили условное название дене-кавказские языки, но они были отнесены С.А. Старостиным к группе современных языков, нерегулярной цепочкой расположенных по Евразии, с заходом их ветви в Америку, в языки американских индейцев. Похоже, что эта цепочка маркирует путь эрбинов по Евразии, а заход в Америку – их родственной (на уровне «дяди») гаплогруппы Q. Поэтому древний язык эрбинов, в динамике его миграции по Евразии, и далее через Кавказ, Месопотамию, Ближний Восток, по Средиземноморью до Пиренейского полуострова и далее по континентальной Европе в период 4800-3000 лет назад, а местами и намного позже (пример – современные баски, язык которых перекликается с северо-кавкаскими языками), был назван языком «эрбин». В конце II тыс. – первой половине I тыс. до н.э. язык эрбин был вытеснен – кроме языка басков и некоторых других не-ИЕ языков древней Европы) – индоевропейскими языками, привнесенных в Европу ранними кельтами, фракийцами, иллирийцами, и с тех пор бóльшая часть Европы заговорила на ИЕ-языках.
Как указывалось выше, в ходе заселения эрбинами Европы между 4800 и 3000 лет назад (и позже) там пропали почти все мужские автохтонные гаплогруппы, включая С, E1b, F, G2a, H2, I1, I2a, I2c, J2, K, R1a, и только R1b интенсивно приумножалась, как приумножались и автохтонные женские, мтДНК. С тех пор гаплогруппа R1b продолжает оставаться доминирующей по численности гаплогруппой центральной и западной Европы, и в настоящее время достигает там 60%. На 95% (или более) это потомки двух пиренейских субкладов R1b-P312 и R1b-U106.
Гаплогруппа R1a, как рассказывалось выше, прошла из Южной Сибири на запад, в Европу, по южной дуге, в отличие от эрбинов, которые прошли на запад сначала по северной дуге, через Северный Казахстан до волжских степей, расселились по Русской равнине в период 8000-4000 тысяч лет назад, до прихода с запада ариев, носителей гаплогруппы R1a. Видимо, последней культурой эрбинов была абашевская (с преобладающими захоронениями на спине) с датировкой 4200-3700 лет назад, которая по времени почти совпадает с фатьяновской культурой (4300-3500 лет назад) средней полосы России (с преобладающими захоронениями в скорченном положении на боку и гаплогруппой R1a в ископаемых образцах ДНК). Первой гаплогруппой, которую можно считать арийской, была R1a-Z645, образовавшаяся в Европе 5500-5200 лет назад, в свою очередь образовавшая субклады Z93, Z283 > Z282 > Z280 (все примерно 5000 лет назад), Z284 и M458 (оба примерно 4700 лет назад), перешедшие на восток, на Русскую равнину. Для R1a-Z284, который получил название «скандинавский субклад», место происхождения менее ясно, или на Русской равнине с последующим переходом его в западную Скандинавию (не в Финляндию, которая технически относится к Скандинавии), или образовался сразу в Западной Скандинавии.
Субклад R1a-Z280 с сопровождающими родственными субкладами образовали археологическую культуру шнуровой керамики (5200-4300 лет назад), которая, возможно, образовалась из культуры шаровидных амфор (5400-4800 лет назад) и/или баденской (5600-4800 лет назад). Место обитания носителей субклада R1a-Z645-Z93 со времени его образования (примерно 5000 лет назад) в течение нескольких последующих веков пока неизвестно, но далее, начиная примерно с 4500 лет назад, он прошел по степным и лесостепным регионам юга теперешних Украины и России, и разошелся на несколько миграционных путей. Уже 4313-4060 лет назад ископаемые носители R1a-Z645 были найдены в синташтинской культуре на Южном Урале (еще один образец – датировка 4141-3911 лет назад), и современные потомки субклада Z93 живут в Индостане сотнями миллионов человек, а также на Алтае, в Китае, на Ближнем Востоке, в странах Персидского залива. Поскольку субклады Z93, Z280 и M458 гаплогруппы R1a образовались из арийского субклада Z645, то все они с полным основанием могут называться арийскими, и не из политических соображений, как некоторые считают, а исходя из исторических данных. Политика здесь совершенно не причем, так как название «арийский» относится только к древним временам, с 5500-5200 лет назад до 2500 лет назад, середины I тыс. до н.э., когда понятие «арийский» было упомянуто в последний раз (насколько нам известно), ахеменидским царем «добронравным» Дарием I (правил в 522-486 гг. до н.э.) на скальном барельефе в Накше-Рустаме в Иране.
Гаплогруппа N образовалась из гаплогруппы NO примерно 36800 лет назад, ее родительская NO образовалась примерно 41500 лет назад, уже позже жизни усть-ишимского человека на Иртыше (см. выше). Происхождение гаплогруппы N относят к юго-восточной Азии, ее носители наиболее многочисленны в Китае, но данные по ископаемым образцам гаплогруппы N там отсутствуют. По современным представлениям, носители гаплогруппы N мигрировали в своей части на север, в алтайский регион, и уже там она, видимо, в значительной степени имела субклад N1a1 (образовался примерно 15500 лет назад). Его сопровождали субклады N1 (образовался 22100 лет назад), N1a и N1b (образовались 18200 лет назад). Их нижестоящие субклады имеют представители коренных сибирских народов, но якуты имеют N1a1. Далее носители N1a1 прошли еще севернее, и перевалили через Уральские горы. Это могло быть в регионе нынешнего Пермского края (отмечено красным цветом на карте ниже), поскольку пермский язык по современной классификации входит в состав финно-угорской ветви уральских языков. Но где бы ни был этот переход, это произошло вряд ли ранее середины II тыс. до н.э., поскольку общий предок современных носителей гаплогруппы N1a1 жил примерно 3200-3500 лет назад (по разным выборкам). Нет никаких указаний, что древние носители гаплогруппы N1a1 на Русской равнине прошли бутылочное горлышко популяции раньше этого времени.
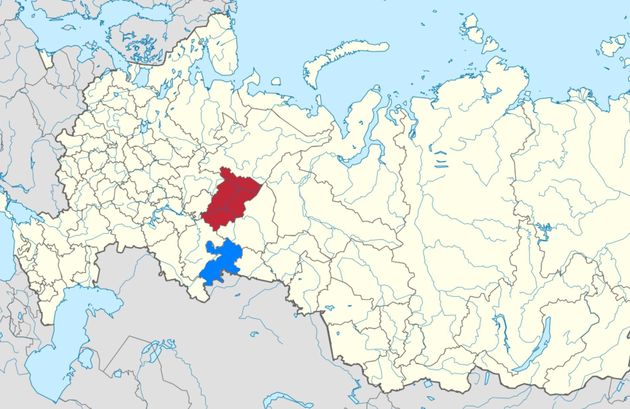
Пермский край (красный цвет) и Челябинская область (синий цвет). В районе нынешнего Пермского края носители гаплогруппы N1a1 могли перейти Урал с сибирской стороны на европейскую 3500-3200 лет назад. В Челябинской области находится синташтинская культура (R1a-Z645) и археологический участок Аркаим (название современное).
После перехода на европейские склоны Уральских гор, или раньше того, носители гаплогруппы N1a1 разошлись на несколько миграционных путей. Предки современных венгров гаплогруппы N ушли на запад, в Паннонию, принесли с собой финно-угорские языки угорской группы, родственные мансийским и хантыйским языкам, языки остались, а сами не выжили, сейчас носителей гаплогруппы N в Венгрии всего 0.5%. Предки современных литовцев, латышей и эстонцев прибыли на берега Южной Балтики примерно 2500 лет назад, в середине I тыс. до н.э., и образовали «балтийский союз» с обитавшими там как минимум уже два тысячелетия носителями гаплогруппы R1a-Z280. С тех пор образовались три перечисленных этноса, в каждом содержание R1a и N1a1 поровну, примерно по 40%, остальное – минорные по численности гаплогруппы R1b, I1, I2a, E1b, T, Q. Предки современных финнов прибыли на территорию нынешней Финляндии 2000-1500 лет назад, в первой половине I тыс. н.э. Структура их гаплотипов существенно отличается в ряде фрагментов от гаплотипов южных балтов. Различаются и языки – у литовцев и латышей языки индоевропейские, у эстонцев и финнов – угро-финские.
Носители гаплогруппы I2a относятся к числу коренных гаплогрупп Европы, во всяком случае после завершения последней ледниковой эпохи (30-10 тысяч лет назад). В ходе заселения Европы эрбинами уцелевшие носители гаплогруппы I2a разбежались на две половины, одна половина оказалась на Британских островах, и имеет общих предков около 5000 лет назад, другая – на Дунае и в Карпатах, с общим предком примерно 2300 лет назад, после прохождения бутылочного горлышка популяции. Последняя группа – в основном славяне, сейчас проживающие от Греции (10% от мужского населения) до Польши (6%), проходя через максимум на Дунае и в Карпатах, например, у боснийских хорват в среднем 71% гаплогруппы I2a.
Итак, выше изложена «приоритетная» гипотеза происхождения основных гаплогрупп, составляющих современные этносы Восточной и Центральной Европы. Посмотрим, как на это описание повлияли недавние данные о сотнях ископаемых ДНК. Скажем сразу, что ни одного из положений «приоритетной гипотезы» данные об ископаемых ДНК не опровергли, хотя, конечно, внесли важные дополнения. Как показано выше, никаких новых данных (ископаемых R1a) на протяженном миграционном пути гаплогруппы R1a от Южной Сибири до Балкан не появилось, хотя в Анатолии и на Балканах ископаемых образцов R1a пока найдено не было. В Анатолии из 15 древнейших (до настоящего времени) образцов ДНК неолита (8400-8200 лет назад) половина (7 образцов) были гаплогруппы G2a (плюс один G), остальные – C1a, H2, J2a, I и I2a. На Балканах образцы R1a тоже пока найдены не были, как описано выше, хотя там проверили всего несколько из сотен захоронений (два образца, один из них женский, из 200 захоронений в Лепенском Вире). Тем не менее, скорченные положения ряда костяков могут свидетельствовать, во всяком случае до проверки, о наличии там мезолитической и неолитической гаплогруппы R1a. Если кто считает, что этих данных недостаточно, пусть предложит обоснованный миграционный путь гаплогруппы R1a в Европу, поддержанный системными данными.
Важным дополнением недавнего времени явились свидетельства о древнейших образцах гаплогруппы R1a-M459 в Карелии (единичный образец с датировкой 7000-7500 лет назад) и в культуре ямочно-гребенчатой керамики в Эстонии R1a-M459-YP1272 (единичный образец с датировкой 5900-3800 лет назад), а также еще более древние субклады R1a-M420 на Украине с датировками 10643, 7100 и 6390±90 лет назад (последний – R1a-M459). Это – немного, но принципиально. Субклады архаичные, таких субкладов с такими датировками общего предка сейчас не обнаружено, точнее, найдено несколько десятков, но с датировкой общего предка 2400-3400 лет назад (33 гаплотипа в 67-маркерном формате субклада R1a-M420-YP4141 дают датировку общего предка 3440±380 лет назад; 18 гаплотипов в 67-маркерном формате субклада R1a-M420-М459-М198 дают датировку общего предка 2360±290 лет назад; 6 гаплотипов в 67-маркерном формате субклада R1a-M420-M459-YP1272 дают датировку общего предка 2500±400 лет назад). Таким образом, следует различать древние образцы ДНК, от которые не осталось современных прямых предков, и образцы, от которых есть современные общие предки. Иначе говоря, генеалогия может терминироваться в древности, а может продолжаться в наших современниках.
Как отмечает статья (Mathieson и др., 2017), скорченное положение костяков очень характерно для мезолита Восточной Украины. С переходом в неолит положение костяков смещается к захоронению на спине. Это может означать смещение мужского населения от R1a в мезолите к R1b в неолите.
Итак, дополнение к «приоритетной гипотезе» может выражаться в том, что после прихода носителей гаплогруппы R1a на Балканы 10000-8000 лет назад они могли продвинуться на Украину, что не так далеко от Балкан, а также на северо-восток Европы с заходом на острова Онежского озера и прилегающие регионы, и через тысячелетия образовать культуру ямочно-гребенчатой керамики. В принципе маршрут мог быть обратного направления, но у нас нет никаких прямых или даже косвенных данных о том, как носители R1a могли попасть на Украину, Балтику и в Карелию, не заходя западнее в Европу. Без данных о таком маршруте дискуссии на этот счет беспочвенны. Опять же, надо в первую очередь исходить из ранее выдвинутой приоритетной гипотезы, и сначала обоснованно отвергнуть ее, и только затем выдвигать новые варианты. Иначе получится безосновательная каша, не имеющая никакой научной ценности.
Вот, пожалуй, и всё, что ископаемые образцы гаплогруппы R1a позволили добавить к приоритетной гипотезе.
Переходим к гаплогруппе R1b. Опять, приоритетная гипотеза, изложенная выше, не претерпела никаких корректировок. Но есть важные дополнения – в нарвской культуре в Латвии найдены два образца гаплогруппы R1b (один из них субклада Р297), с датировками 7690±100 и 7030±225 лет назад. Субклад Р297 является дочерним по отношению к субкладу R1b-L278 хвалынской культуры, и, как подсказывают нам археологи, его носители могли подняться по Волге и последующим речным путям до Балтики. Помимо того, в Латвии была найдена еще большая серия из восьми древних образцов R1b-Р297, с датировками от 9220 до 7500 лет назад, и один образец с датировкой 6060 лет назад, все восемь образцов имели тот же субклад, что и в нарвской культуре.
Тот же субклад, что и в хвалынской культуре, R1b-L278, был найден на Украине в пяти ископаемых образцах ДНК, и еще в двух образцах R1b-L278-L754, дочерним к первым двум (датировка всех четырех примерно 7100 лет назад), причем захоронения были в основном на спине, что характерно для погребального обряда носителей гаплогруппы R1b. Помимо того, древние (архаичные) образцы R1b были найдены в Болгарии (L278, 6448 лет назад), в Румынии (L278, 9335 лет назад), в Германии (L278-L754, 5541 лет назад), Испании (L278-L754, 7130 лет назад), в Лепенском Вире (L278-L754, 8017 лет назад). Самый древний образец R1b был найден в Италии (Виллабруна, L278-L754, 13980 лет назад).
Эти дополнения действительно очень важные – приоритетная гипотеза рассматривала только миграции в Европу эрбинов бронзового века, с их прибытием на Пиренейский полуостров примерно 4800 лет назад, и дальнейшее заселение Европы как культуры колоколовидных кубков. Ископаемые образцы показали, что эрбины прибыли в Европу значительно ранее, сразу после завершения последней ледниковой эпохи (10 тысяч лет назад), правда, они имели архаичные субклады, L278-L754, это – второй и третий уровень после исходной гаплогруппы R1b-M343:
R1b-M343 > L278 > L754
Эти субклады практически не дожили до настоящего времени, поскольку современные носители гаплогруппы R1b происходят от предков только десятого уровня, которые и образовались на Пиренеях примерно 4800 лет назад (ниже отмечены жирным шрифтом).
R1b-M343 > L278 > L754 > L388 > P297 > M269 > L23 > L51 > L151 > P312, U106
Естественно, первая приоритетная гипотеза, которая строилась на гаплотипах наших современников, не могла предусмотреть те древние R1b, носители которых давно вымерли.
Относительно гаплогруппы N1a1 – ископаемые образцы ДНК только подтвердили первую приоритетную гипотезу тем, что в культуре ямочно-гребенчатой керамики (ЯГК) обнаружена гаплогруппа R1a, а не N. Образец единичный, но показательный, тем более что те же архаичные субклады обнаружены на Украине и в Карелии. Подтвердили – потому что по приоритетной гипотезе носители гаплогруппы N1a1 появились на Русской равнине не ранее 3200-3500 лет назад, и находка их в культуре ЯГК серьезно модифицировала бы гипотезу. Это уже была бы модификация, а не дополнение. Справедливости ради надо отметить, что по сообщению Чекуновой и др. (2014) на северо-западе России, в Смоленской области, на территории жижицкой археологической культуры с принимаемой датировкой середины III тыс. до н.э. найден образец N1a1. Сам образец не датировался, описание дано в виде краткого сообщения, в части самого образца – буквально в две строки, археологического контекста не дано, работа с древними образцами описана только для мтДНК, N1a1 описан, конечно, как «финно-угорский», а древние носители – «финскими», как и то, что этот образец отнесли к «распространению культуры ямочно-гребенчатой керамики», что типично для попгенетиков. Достоверность типирования и датировки самого образца (которой, повторяю, не было) проверить невозможно, тем более что он мог попасть туда в более позднее время. Итог – единичный образец в таком неясном описании можно только иметь в виду при наличии перекрестных проверок в будущем.
Общий итог – по всему списку ископаемых гаплогрупп Y-хромосомы из 126 образцов (Allentoft и др., 2015) и 270 образцов (Mathieson и др., 2017) гаплогруппа N (и субклады) не присутствовала.
Относительно гаплогруппы I2a «приоритетная гипотеза» сообщает, что их носители были одними из коренных жителей Европы, вместе с носителями гаплогрупп С, E1b, F, G2a, H2, I, I1, I2c, J2, K, R1a, которые пропали из Европы в ходе заселения эрбинами, носителями гаплогруппы R1b, в основном насельниками культуры колоколовидных кубков. Эта культура вышла с Пиреней вместе с носителями субкладов P312 и U106, оба образовались примерно 4800 лет назад (Klyosov, 2010, 2012; https://www.yfull.com/tree/R1b/). Гаплогруппа I2a частью бежала на Британские острова (общий предок современных носителей I2a жил там около 5000 лет назад), частью на Дунай и в Карпаты (общий предок жил примерно 2300 лет назад). Ископаемые образцы согласуются с «приоритетной гипотезой» в том отношении, что гаплогруппа I в Европе в большинстве случаев найдена в древних захоронениях, с датировками от 12960 лет назад (Франция) и 10040 лет назад (Сербия), хотя первый случай удивителен, на территории Франции 13 тысяч лет назад должен был быть мощный ледник, и до 4500 лет назад. Это – 16 образцов из 21 найденных, и эти образцы были найдены по всей Европе – Балканы, Италия, Франция, Германия, Украина. Остальные пять образцов были найдены с датировками после 4000 лет назад, то есть уже к концу предполагаемого истребления коренных жителей Европы, из этих образцов три в Швеции (3800, 3300 и 3200 лет назад), один в Италии (3900 лет назад) и один в Венгрии (3970 лет назад). После этого ископаемых I уже не находили. Пока, во всяком случае.
Аналогичная ситуация – с гаплогруппой I2a. Из 46 ископаемых образцов I2a сорок два, то есть 91%, найдены в древнейших захоронениях, с датировками от 10074 лет назад (на Украине) до 4500 лет назад, причем преимущественно более 4900 лет назад (от 4900 до 4500 лет назад найдены только три образца – в Испании, Болгарии и на Украине). Остальные четыре образца, включая один на территории ямной культуры в Калмыкии, с датировкой 4446 лет назад, были найдены в Испании (4407 лет назад) и Венгрии (3700 и 3600 лет назад). После этого ископаемых I2а уже не находили. По «приоритетной гипотезе» их и не должно было быть, во всяком случае много, они проходили бутылочное горлышко популяции до примерно 2300 лет назад, то есть почти до конца старой эры.
Похожая ситуация и с носителями гаплогруппы I2c, их найдено всего три образца из нескольких сотен, два самых древних, с датировками 8300 и 7600 лет назад (в Турции и Швеции), и один, намного более поздний, в Германии, с датировкой 3990 лет назад. Это тоже не противоречит «приоритетной гипотезе», согласно которой автохтонное мужское население Европы было истреблено в период 4800-3000 лет назад. Естественно, к завершению этого периода число их захоронений прогрессивно уменьшалось.
Итак, повторяем основной вывод – новые данные по ископаемым ДНК важны для уточнения датировок древних миграций, как и для уточнений и дополнений выдвигаемых ранее «приоритетных гипотез», основанных на ДНК-генеалогическом анализе гаплогрупп, субкладов и гаплотипов. Но в большей части оказалось, что ископаемые ДНК с их гаплогруппами и субкладами понадобились главным образом для описаний, какие гаплогруппы и субклады найдены, и какие там археологические датировки. Выработанную ранее картину это, как правило, не меняет, хотя в некоторых случаях дополняет. Иногда это важные дополнения, расширяюшие наши знания, но в большей части не очень важные. Видимо, основная причина такого «не очень» заключается в том, что ископаемые образцы в подавляющем большинстве случаев относятся к Западной, Центральной и Восточной Европе, то есть к тем регионам, для которых уже определены десятки и сотни тысяч современных гаплотипов и их субкладов, и древняя ДНК-генеалогическая картина уже в целом ясна. Ископаемые образцы ДНК просто подтверждают во многих случаях наши представления о бутылочных горлышках популяций. Действительно большое продвижение в наших знаниях было бы, если бы палеогенетики определяли гаплотипы в ископаемых образцах ДНК. К сожалению, они этого в принципе не делают. Вместо того они занимаются интерпретационными фантазиями, причем, как правило, в вязком, неопределенном, гадательном виде, с постоянными ошибками в тех редких случаях, когда они осмеливаются делать какие-то более или менее определенные выводы. Это – воистину трагедия современной «широкогеномной науки».
Анатолий Алексеевич Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















