
Читатели Переформата, которых интересуют не только исторические и публицистические статьи, но и материалы по ДНК-генеалогии, наверное, обратили внимание, что в начале 2016 года сайт Академии ДНК-генеалогии пополнился разделом «Гаплокарты». Таким, пока еще редко употребляющимся сокращением, автор этих строк назвал карты, на которых приведена статистика по основным Y-хромосомным гаплогруппам Евразии. Идея нанести на карту такие данные появилась, очевидно, с того самого момента, как они начали поступать в более-менее значимых количествах, то есть с конца 1990-х – начала 2000-х годов. Сейчас в книгах, журналах и Сети можно найти огромное количество карт на любой вкус и цвет, что в той или иной мере касаются Y-ДНК. Казалось бы, что нового может дать очередная их версия? Ответу на этот вопрос посвящен краткий обзор карт и таблиц, которые размещены в указанном разделе.
Вначале следует объяснить, чем не устраивают существующие карты. Прежде всего, при всем разнообразии, среди них нет таких, что в полной мере могли бы дать наиболее полный обзор распределения Y-гаплогрупп по регионам и этническим группам, где привлекались бы последние данные с максимально доступной на сегодняшний день репрезентативностью и разрешением по филогении. Имеющиеся в наличии карты и таблицы такой связной картины не дают. Они либо ограничиваются какой-либо конкретной гаплогруппой и регионом, либо используют устаревшую номенклатуру, имеют недостаточное разрешение по филогении, а зачастую опираются на выборки с низкой статистической значимостью. Например, таблица, размещенная в Википедии, предлагает информацию об Y-гаплогруппах голландцев по выборке из 27 (sic!) гаплотипов, со ссылками на публикации 2000 и 2004 гг. По миллиардным Индии и Китаю нет даже этого. Альтернативой подобной попгенетической архаике можно было бы считать карты и таблицы с проектов FTDNA, но их составители зачастую бросаются в другую крайность – настолько сильно дробят участников на категории и подкатегории, что для восстановления общей картины по тому или иному этносу требуется приложить немало усилий.
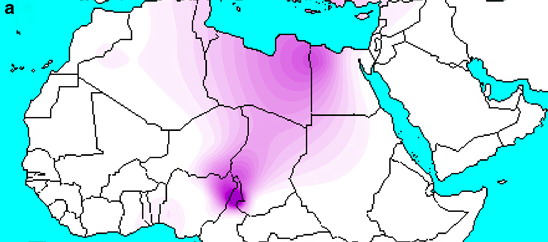
Наконец, нельзя не обойти вниманием, что часто приводимые в справочной литературе карты гаплогрупп, имитирующие карты плотности населения, грешат неоправданной детализацией, не соответствующей реальному уровню знания. Это почти точная аналогия попгенетическим датировкам, что рассчитываются с погрешностями в десятки процентов, но с точностью до четвертого знака после запятой. Бывают и откровенные курьезы, как, например, шлейф ветви R1b-V88, проходящий через одно из самых малонаселенных мест на Земле – нагорья Тибести и Энеди в Сахаре (см. ниже). Тот, кто знает, как работают компьютерные программы, рисующие такие карты, понимает, что это издержки алгоритма, но многие принимают его причуды за чистую монету и устраивают бурное обсуждение по поводу того или иного максимума на карте плотности.
Итак, чтобы пройти между Сциллой скудости исходных данных и Харибдой сомнительной детализации, требуется корректная постановка задачи и ясное понимание того, насколько полную информацию могут предоставить существующие данные по Y-хромосомным гаплогруппам. Чтобы добиться поставленной цели, то есть дать общий и, по возможности, объективный обзор с максимально возможным охватом по географии и филогении, следует, во-первых, отказаться от чрезмерной детализации и наносить на карты только статистику по ключевым гаплогруппам с их привязкой по странам или регионам больших стран. Тем самым можно получить сопоставимые данные по всем доступным регионам, без перекоса в пользу более изученных. Излишняя перегруженность карты деталями, помимо того, делает ее трудной для восприятия. От идеи представить карты в виде контуров плотности, при всей их наглядности, также следует отойти, чтобы не возникало соблазна интерпретировать «фантомы», созданные компьютерными программами. Самой надежной и наглядной формой подачи на данный момент можно считать хорошо проверенные круговые диаграммы, размещенные в соответствующих местах.
Следующим этапом, после формы представления, следует считать отбор исходных статистических данных. Во-первых, они должны быть репрезентативны, то есть отображать реальную статистику по тому или иному региону или этнической группе, без серьезных перекосов. Во-вторых, они должны содержать информацию по всем гаплогруппам, задействованным в обзоре, либо должны позволять извлечь такую информацию из гаплотипов, не типировавшихся на снипы. И, в-третьих, желательно иметь как можно более свежие данные, чтобы избежать ошибок типирования, что были нередки на начальном этапе сбора информации. Как оказалось, значительная часть материала, собранного и опубликованного в самых цитируемых работах попгенетиков, не проходит по какому-либо из этих критериев, а то и по всем сразу. Этих недостатков лишены данные с географических ДНК-проектов, опубликованные на сайте компании FTDNA, но их охват по регионам очень неравномерен. Например, по Азии репрезентативные данные имеются только для армян, грузин и арабов из стран Персидского залива. Недостающие данные можно получить из судебно-медицинских выборок, опубликованных в специализированных журналах, а также в базе данных YHRD. Хотя в них, как правило, не определяют снипы, отнесение можно с достаточно высокой степенью надежности оценить по гаплотипам в 17- и 23-маркерных форматах, принятых экспертами-криминалистами в качестве стандарта. Методика работы с этими наборами хорошо отработана, и ошибки типирования в последние 3-4 года стали большой редкостью. В ряде случаев, этим данным отдавалось предпочтение даже по некоторым странам, хорошо представленным в проектах FTDNA, если требовалась привязка к конкретным регионам. Например, сводная статистика по русским была взята из полевой выборки 2008 года, чтобы иметь представление о географии гаплогрупп в пределах России. Ссылки на оригинальные работы и данные в табличной форме по каждой из диаграмм приведены в таблицах, размещенных рядом с картами.
Пришлось пока отказаться от картирования Индии, точнее ее самой населенной северной части, потому что имеющиеся данные, хотя и исчисляются сотнями гаплотипов, не дают желаемой связной картины. В публикациях в обилии представлены различные этнические и социальные группы индийцев, но без соотнесения с демографией. Насколько они отображают ситуацию в целом по региону, судить сложно. К примеру, можно ли считать репрезентативной выборку русских, если она состоит из 50 человек с фамилией «Иванов», 50 – с фамилией «Клёсов» и 50 – с фамилией «Рожанский»? Может быть, да, а может быть, нет. Без отдельной проверки вывод сделать нельзя, что мы и наблюдаем с индийцами. Выборки, собранные без дробления на касты и племена, пока имеются только по тамилам с юга Индии и мусульманам из Бангладеш и Пакистана. По другим странам Азии таких проблем не возникало. Насколько можно судить, информация по гаплогруппам Азии в таком объеме публикуется впервые. Карты для Африки, Океании и коренного населения Америки пока не готовились из-за недостатка данных, а также из-за сравнительно небольшого набора гаплогрупп, присутствующих у живущих там народов.
На этом первую часть обзора, а именно постановку задачи и описание исходных данных, можно считать завершенной. Во второй части предлагаю краткую характеристику генеалогических линий, представленных на гаплокартах.
Встречается крайне редко, а потому включена в раздел «прочие. Тем не менее, в Западной Европе живут носители загадочной гаплогруппы А1а, которой была посвящена заметка на Переформате. Гаплогруппа A1b найдена у арабов Персидского залива и Ливана, где представлена ветвью A1b1b-М32, пик распространения которой приходится на Эфиопию и нилотские народы Восточной Африки. Помимо арабов, единичные гаплотипы той же ветви отмечены у армян, в Казахстане, на Русской равнине и Британских островах. Очевидно, за пределы Восточной Африки субклад М32 распространился как минорная ветвь в миграциях народов – носителей галогруппы E1b1b1-М35, со стороны Африканского Рога, в том числе и путем работорговли.
Также из категории «прочие». Встречается за пределами Африки реже, чем гаплогруппа А. Обнаружена у арабов и хазарейцев из Афганистана. Вероятно, маркирует потомков рабов, вывезенных арабскими купцами из Восточной Африки. Следует отметить, что рабство в странах Ближнего Востока отличалось от рабства в Новом Свете XVI-XIX веков тем, что в нем не было столь выраженной сегрегации между рабами и хозяевами. В течение нескольких поколений потомки раба, если у него была семья, становились равноправными членами сообщества, поскольку ислам в его самой строгой форме отрицает сословное деление для его последователей. При удачном стечении обстоятельств человек, некогда купленный на невольничьем рынке, мог достичь самого высокого социального статуса, как показывают примеры мамлюкских султанов в Египте и Палестине.
Одна из древнейших и самых разветвленных гаплогрупп. Присутствует повсеместно, кроме Африки. Найдена у европейцев эпохи палеолита (Костёнки), мезолита (Испания) и раннего неолита (Венгрия), но в настоящее время ее в Европе настолько мало, что в выборке европейцев она входит в категорию «прочие». В Центральной и Восточной Азии, напротив, это одна из основных гаплогрупп, что представлена почти исключительно субкладом С2-М217, который расходится на большое количество ветвей с общим предком, жившим около 34000 лет назад. Родственный ему, и более древний «костенковский» субклад С1-F3393 рассеян от Европы до Океании. В выборках, представленных на гаплокартах, его ветви в значимых количествах присутствуют у японцев и филиппинцев на уровне 3-5%, а также найдены в древней ДНК из Красноярского края и с северо-востока Китая.
Является предковой к гаплогруппам D и E, характеризуется цепочкой в 40 снипов в формате YFull. Единственный подтвержденный представитель – араб из Сирии. Сообщения из ранних статей о находках DE в Африке и на Тибете пока сложно подтвердить или опровергнуть из-за высокой вероятности «недотипирования», что часто имело место на начальном этапе сбора информации по Y-ДНК. Возможно, среди нескольких тысяч гаплотипов из полевых выборок Китая и Юго-Восточной Азии есть вероятность найти представителей чрезвычайно редких линий, восходящих непосредственно к DE, но из-за отсутствия данных по снипам их пока невозможно распознать.
Практически не встречается за пределами Восточной и Юго-Восточной Азии, где является одной из принципиальных генеалогических линий у тибетцев и японцев. Расходится на несколько далеко отстоящих ветвей с общим предком, жившим около 46000 лет назад, из которых субклад D-M64.1 ограничен почти исключительно Японским архипелагом и восходит, очевидно, к его древним обитателям из культуры Дзёмон (шнуровой керамики, в переводе с японского). Большой перевес гаплогруппы D на Тибете создается за счет одной из ветвей субклада D-P47, не имеющей пока своего снипа, но легко узнаваемой по редкому значению DYS392=7. Ее предок жил сравнительно недавно – около 4800 лет назад. Та же ветвь присутствует и у других народов Восточной Азии, но намного реже. Остальные ветви гаплогруппы D встречаются очень редко, и в отсутствие данных по снипам ее носителей распознать весьма сложно.
Наряду с J, O и R, входит в четверку «супер-гаплогрупп», число носителей которых исчисляется сотнями миллионов, а география захватывает целые континенты. Для гаплогруппы Е этот континент – Африка, хотя из рассмотрения гаплогрупп DE и D следует, что ее корни находятся вне Африки. На Черном Континенте представлены все основные субклады этой древней (около 54000 лет до предка) гаплогруппы, из которых только один, E1b1b1-M35, вышел за его пределами в значимых количествах. Соответственно, на гаплокартах отмечена статистика только по М35, а остальные внесены в категорию «прочие». Субклад E-M35 в Евразии расходится на 2 основные ветви, Е-М78 и Е-Z287, что имеют «возраст» около 10000 и 12000 лет, соответственно. В свою очередь, они дробятся на большое количество дочерних ветвей, из которых можно выделить E-V13, специфическую для Европы и начавшую свой рост около 3700 лет назад, как многие другие генеалогические линии Европы, прошедшей через демографический кризис на рубеже III и II тысячелетий до н.э. С другой стороны, находки ископаемых гаплотипов Е-V13 и E-M78 с датировками около 7000 лет назад в Испании и Венгрии говорят о том, что представители этих ветвей стали заселять Европу еще со времен раннего неолита.
Подтвержденных носителей этой гаплогруппы, не относящихся к дочерним GHIJK, в настоящее время известно крайне мало. Это единичные гаплотипы из Южной и Юго-Восточной Азии, к которым, возможно, добавится какое-то количество гаплотипов из выборок по этим регионам, не типированных пока на снипы. Для программы-предиктора они неотличимы от тех, что принадлежат к гаплогруппам G или H. Сопоставимое количество носителей гаплогруппы F было найдено в ископаемой ДНК в Европе, в том числе в останках 39000-летней давности из Румынии. Такой результат вполне ожидаем, если принять во внимание, что в гаплогруппы, дочерние к F, входит подавляющее большинство современных жителей Евразии, Океании и Америки.
Современные представители этой гаплогруппы имеют общего предка, жившего около 25000 лет назад, что заметно меньше, чем у рассмотренных выше линий. На гаплокарте дана статистика по самому населенному ее субкладу G2a-P15, что имеет «возраст» около 17000 лет и рассеян от Западной Европы до Центральной Азии. Несколько лет назад большим сюрпризом стали находки этой гаплогруппы в ископаемой ДНК Европы, где сейчас доля G2a невелика. В настоящее время, когда число образцов ископаемой Y-ДНК перевалило за 100, можно считать вполне установленным фактом то, что это была одна из основных линий Европы эпохи неолита. О распределении G2a в Азии той же эпохи пока нет информации, но, судя по современным данным с Кавказа и из Ирана, ее представители жили и там. Статистика по другим, меньшим по численности и более локальным по своей географии ветвям гаплогруппы G, собрана в категории «прочие». Это субклады G1 и G2b, рассеянные с низкой частотой в Передней и Средней Азии. Обе эти редкие ветви найдены также у евреев-ашкенази, что предполагает присутствие гаплогрупп G1 и G2b на Ближнем Востоке до начала диаспоры евреев.
Несмотря на раннее время расхождения на ветви (около 48000 лет до предка), эта гаплогруппа имеет довольно компактный ареал, ограниченный в основном Южной Азией. В Европе встречается, как правило, у цыган и их потомков, принесшим ее из Индии. Однако в ископаемой ДНК из Европы времен неолита были обнаружены представители ныне крайне редкой гаплогруппы Н2-Р96, что предполагает более широкую географию Н в прошлом.
Находится в родстве с распространенной на Ближнем Востоке и в Средиземноморье гаплогруппой J, с которой образует сводную гаплогруппу IJ, характеризуемую цепочкой из 64 снипов в формате YFull, но связана с Европой, где, очевидно, зародилась и начала расходиться около 27000 лет назад. В Азии встречается очень редко, а потому отнесена на гаплокарте Азии в категорию «прочие». На гаплокарте Европы, в силу особой значимости в этногенезе народов Европы, дана статистика по трем принципиальным дочерним гаплогруппам: I1-M253, I2a1-P37.2 и I2a2-M436. На сегодняшний день пока не найдено ни одного надежно подтвержденного гаплотипа из родительской гаплогруппы I, в том числе и в ископаемой ДНК. Перечисленные выше дочерние гаплогруппы охватывают абсолютное большинство современных носителей гаплогруппы I, за исключением очень редких I2b-L415 и I2c-L596, что помещены в категорию «прочие». Одна из ветвей I2c, а именно Y16419, компактно располагается в Восточном Причерноморье, где ее доля доходит до 5% в выборке по Восточной Грузии. Сравнительно небольшое, около 3500 лет назад, время жизни ее предка и находки родственных гаплотипов в Европе эпохи мезолита и раннего неолита дают основание считать, что эта линия имеет европейское происхождение.
Долгое время для этой гаплогруппы, рост которой начался около 4300 лет назад, постулировали североевропейское или даже скандинавское происхождение. Однако из гаплокарты Европы следует, что ее география намного шире, и охватывает весь континент, за исключением юга Балканского и Апеннинского полуостровов. Самые рано отошедшие ветви I1 распространены в Центральной и Восточной Европе, но почти не захватывают Скандинавский полуостров, что мало согласуется со скандинавской гипотезой. Находка I1 в ископаемой ДНК из Венгрии времен неолита (культура линейно-ленточной керамики) также можно считать доводом в пользу континентального происхождения этой гаплогруппы. Однако пока нет возможности установить, в каком месте Европы и в составе каких народов начался ее рост после прохождения бутылочного горлышка на рубеже III и II тысячелетий до н.э.
История этой гаплогруппы уходит ко временам палеолита (около 18500 лет до предка), но в настоящее время подавляющее большинство ее носителей принадлежит к ветви CTS10228, рост которой начался в эпоху античности, около 2300 лет назад. Высокий численный перевес и значительный процент этой ветви среди народов юго-востока Европы создали иллюзию балканского происхождения всей гаплогруппы I2, что, судя по ее филогении и данным древней ДНК, вряд ли соответствует действительности. Все остальные ветви гаплогруппы I2a1, кроме уже упомянутой CTS10228, рассеяны по Западной Европе. Одна из них, PF4188, является основной генеалогической линией жителей Сардинии, но, как и в случае с восточноевропейской ветвью CTS10228, ее предок жил сравнительно недавно – на рубеже нашей эры. В настоящее время пока сложно сказать с определенностью, где зародилась гаплогруппа I2a1 и каковы были ее пути при заселении Европы.
Время жизни предка I2a2 совпадает с тем, что оценено для I2a1, но, в отличие от последней, в ней нет таких многолюдных линий, как CTS10228. Она расходится на несколько примерно равных по численности ветвей, доля которых достигает 6-7% в выборках из Дании, Нидерландов и Англии. Наряду с I2a1 и I2c, носителей этой гаплогруппы идентифицировали в древней ДНК из различных частей Европы. Несмотря на небольшую численность, гаплогруппа I2a2 имеет широкий ареал, и в качестве минорной линии присутствует у всех народов Русской равнины, доходя до Афганистана и уйгуров из оазисов Синьцзяна. Возможно, какая-то часть ее носителей приняла участие в миграциях ариев, но по численности они сильно уступали представителям гаплогруппы R1a, и большинство их линий пресеклось.
Следующая из гаплогрупп-«тяжеловесов», как правило, сейчас редко рассматривается как единое целое. В базах данных и публикациях отдельно идет статистика по дочерним гаплогруппам J1 и J2. В современных выборках пока не найдено гаплотипов, которые можно считать подтвержденными представителями родительской ветви J. По данным древней ДНК, возможно, к ней принадлежал человек, живший около 7200 лет назад на территории современной Карелии, но из-за плохой сохранности ДНК многие ее фрагменты не прочитаны, что оставляет возможность отнесения либо к J1, либо к J2.
Является одной из основных генеалогических линий на Ближнем Востоке и на Кавказе, где представлена преимущественно субкладами YSC0000234 (около 4600 лет до предка) и Z1842 (около 6000 лет до предка), соответственно. Другие ветви встречаются заметно реже, но довольно сильно разбросаны по географии. К одной из них, F6304, вероятно, принадлежал человек, живший на территории Грузии около 13200 лет назад. Судя по этим данным, а также исходя из времени жизни предка всех современных представителей J1 (около 18500 лет назад), эта гаплогруппа с самых ранних времен занимала территории к югу от Большого Кавказского хребта, но более точно локализовать ее прародину пока не удается. В популярных изданиях гаплогруппу J1 нередко называют основной у евреев, но это не совсем верно. В репрезентативной выборке евреев-ашкенази она находится на третьем месте, немного уступая гаплогруппам E и J2.
Одна из самых старых (около 28000 лет до предка) и самых разветвленных генеалогических линий Евразии. Проходит широкой полосой от Пиренейского полуострова на западе до верховьев Хуанхэ на востоке, составляя значительную долю у народов Средиземноморья, Кавказа, Передней и Южной Азии. Многие из дочерних ветвей J2 имеют выраженную привязку к тем или иным регионам и этносам, что позволяет возводить их к миграциям разных эпох, от неолита до раннего Средневековья. Находки ископаемой ДНК из гаплогруппы J2 пока не очень многочисленны, но по ним можно судить о ее широкой географии уже в древние времена. Это Грузия 9600 лет назад, Турция 8400 лет назад, Венгрия 7000 и 3200 лет назад, Армения 3100 лет назад и Алтай начала нашей эры.
Подобно гаплогруппе F, в Евразии найдено очень мало подтвержденных представителей К, кто не входит в дочерние сводные гаплогруппы LT и NOP. Однако значительное количество таких линий существует в Австралии и Океании. В их числе минорные гаплогруппы M и S, с выделением которых в самостоятельные таксономические единицы явно поторопились. Можно ожидать, что в представленных на гаплокарте выборках из примыкающей к Океании Юго-Восточной Азии обнаружатся новые представители гаплогруппы К, но в отсутствие данных по снипам их распознать невозможно, и они, скорее всего, оказались на настоящий момент в списках гаплогрупп N, O или Q. В данных по ископаемой ДНК имеется пока всего один представитель предковой гаплогруппы К, но это самый древний на сегодняшний день человек, у которого была расшифрована Y-хромосома. Он жил в Западной Сибири около 47000 лет назад, а его останки (бедренную кость) нашли близ впадения Ишима в Иртыш, откуда и знакомое многим имя – человек из Усть-Ишима. По сумме косвенных данных, родиной гаплогруппы К можно считать Центральную, Восточную или Юго-Восточную Азию, но локализовать ее более точно пока сложно.
Входит в сводную гаплогруппу LT, в составе которой имеет 32 общих снипа с гаплогруппой T в формате YFull. Время жизни предка современных представителей гаплогруппы L оценивается как примерно 23000 лет назад. В выборках из Южной Азии ее доля достигает 7-15%, в остальных регионах встречается редко, потому на гаплокарте Европы помещена в категорию «прочие». Локальный максимум гаплогруппы L отмечен среди нескольких народов Кавказа. Он достигается за счет молодых ветвей Y6247 (около 1000 лет до предка) и PH8 (около 2400 лет до предка).
Находится в близком родстве с гаплогруппой О, основной генеалогической линией Восточной Азии. Их общий предок из сводной гаплогруппы NO жил около 36500 лет назад. Так же, как и у жившего 15000 годами спустя предка современных представителей гаплогруппы N, их родину можно достаточно надежно поместить в Восточную Азию. В выборках из Китая и Монголии гаплогруппа N занимает третье место по численности, после О и С, а в древней ДНК времен неолита и бронзы с севера Китая она оказывается основной. Среди народов Дальнего Востока и Восточной Сибири, не представленных пока на гаплокарте, гаплогруппа N также одна из основных. В Западной Сибири и в Европе из всех ее ветвей присутствует почти исключительно довольно молодая (около 4500 лет до предка) ветвь L1026, появление которой связывают с распространением финно-угорских языков. В отличие от Китая, в Европе пока мало находок ископаемой ДНК из гаплогруппы N. Это два образца с Валдайской возвышенности (4500 и 1100 лет назад) и один из Венгрии (2900 лет назад).
Видимо, занимает первое место в мире по численности из всех гаплогрупп, поскольку доминирует в густонаселенных странах Восточной и Юго-Восточной Азии. Расходится на большое количество ветвей, филогения и география которых находятся в процессе уточнения. По этой причине, а также чтобы не перегружать карту, на гаплокарте Азии приведена статистика по всей гаплогруппе, без разделения на субклады. Помимо Восточной Азии, гаплогруппа О составляет значительную долю у народов, говорящих на языках австронезийской семьи и расселившихся от Мадагаскара до Гавайских островов. На севере Китая найдено большое количество древней ДНК из различных ветвей гаплогруппы О, но, в отличие от современного населения, по числу находок она уступает гаплогруппам N и Q. Это дает дополнительный аргумент в пользу того, что первоначальный ареал гаплогруппы О охватывал юг современного Китая, а ее современное географическое распределение – результат миграций, начавшихся в эпоху неолита.
Поскольку пока нет подтвержденных сведений о гаплотипах из родительской гаплогруппы Р, обзор продолжает гаплогруппа, которую можно считать безоговорочным лидером по распространению до эпохи Великих географических открытий. Ее ареал к началу Нового Времени занимал все северное полушарие, за исключением Африки, а также Южную Америку. Самое северное (Гренландия) и самое южное (Огненная Земля) места на Земле, где имеется постоянное население, были освоены представителями гаплогруппы Q. Однако, за исключением коренных жителей Америки и нескольких народов Сибири, гаплогруппа Q составляет незначительную долю у жителей Евразии. По абсолютной численности в Старом Свете, очевидно, лидирует ветвь М120, составляющая 2-4% в выборках ханьцев и найденная в древней ДНК на севере Китая. Больше всего различных линий этой очень разветвленной гаплогруппы отмечено в полосе от Алтая до Афганистана, в том числе и в ископаемой ДНК. Видимо, где-то в этом регионе около 32000 лет назад и жил ее предок. Локальные максимумы за пределами Центральной Азии отмечены у евреев-ашкенази (ветвь Y2200, около 1400 лет до предка), чеченцев (YP4000, 1200 лет) и жителей Скандинавии (L527 и L804, 2300-2100 лет), что косвенно говорит о некогда большой мобильности народов – носителей гаплогруппы Q.
Последняя по очереди и самая молодая из рассмотренных «супер-гаплогрупп». Ее предок жил около 31000 лет назад, очевидно, в Азии, как можно заключить из ее родства с азиатскими гаплогруппами NO и Q, а также из находки древней ДНК с палеолитической стоянки Мальта в Прибайкалье, датируемой 24000 годами назад. Следовательно, распространение гаплогруппы R, к началу Нового времени занимавшей Европу, Анатолию и Южную Азию, – результат древних миграций с востока на запад, детали которых пока неизвестны. Современные представители гаплогруппы R принадлежат к одной из трех дочерних гаплогрупп, R1a-M420, R1b-M343 и R2-M479, статистика по которым приведена на гаплокартах отдельно.
Предок этой многомиллионной гаплогруппы жил во времена палеолита, около 18000 лет назад, но абсолютное большинство современных представителей R1a ведет свой род от двух человек, живших намного позднее – около 5500 лет назад, когда начался рост субкладов Z282 и Z93. В свою очередь, они разошлись на многочисленные дочерние ветви, каждая со своей характерной географией и этническим составом. В общих чертах с ними можно ознакомиться по гаплокарте R1a, где указано распределение основных субкладов там, где позволял имеющийся материал. Гаплогруппа R1a найдена в многочисленных образцах древней ДНК, как правило, из археологических культур, связываемых с распространением индоевропейских языков – от культуры боевых топоров на западе до андроновской на востоке. След миграций древних ариев просматривается вплоть до Великой Китайской равнины и Таиланда, а на территории Индии их потомками является до 1/3 населения, преимущественно из ветвей L657 и Z2123. Очень редкие, рано отошедшие ветви YP4141, YP1272, YP1051 и L664 найдены пока в Европе и на Ближнем Востоке, в том числе ископаемый гаплотип из Карелии с датировкой около 7200 лет назад.
Датировки как предка всей гаплогруппы, так и доминирующих субкладов L51 и Z2103 практически совпадают с теми, что найдены в родственной гаплогруппе R1a. Однако ареал гаплогруппы R1b оказывается более разорванным, как можно заключить из составленной для нее гаплокарты. Западноевропейский субклад L51 и «анатолийский» Z2103 перекрываются в выборках из Центральной Европы и Южной Италии, но это, скорее, результат встречного движения с разных сторон, чем свидетельство центрально-европейского происхождения родительской для них ветви М269. Находки древней ДНК во многом соответствуют современному распределению этих субкладов (L51 – запад, Z2103 – восток), но, в отличие от заволжских степей (ямная и полтавкинская археологические культуры), носители гаплогруппы R1b появляются в Западной и Центральной Европе только в середине III тысячелетия до н.э. в захоронениях, относимых к культуре колоколовидных кубков. В пользу сравнительно позднего появления R1b в Европе косвенно свидетельствует и география рано отошедших ветвей V88 и M70, рассеянных по Центральной Азии, Ближнему Востоку и Центральной Африке (V88, см. рисунок выше). В отличие от родственной гаплогруппы R1a, пока не найдено каких-либо древних европейских ветвей R1b, причем на коммерческих выборках в десятки тысяч образцов в высоком разрешении.
Находится в тени более успешной в демографическом плане гаплогруппы R1, в том числе и потому, что ее ареал ограничен в основном Центральной и Южной Азией. Однако даже в Индии и Афганистане ее доля сравнительно невелика, и не превышает 10%. Восходит к предку, жившему около 11000 лет назад, видимо, в Центральной Азии, где живут представители самых рано отошедших ветвей R2. Вопреки распространенному мнению, гаплогруппа R2 у европейских цыган встречается очень редко, лишь в нескольких этнических группах. Родословные немногочисленных европейских представителей этой гаплогруппы восходят не столько к цыганам, сколько к евреям-ашкенази или потомкам уроженцев Ближнего Востока, разными путями попадавших в Европу.
Входит в сводную гаплогруппу LT, но, в отличие от L, не имеет выраженной привязки к каким-либо регионам или этническим группам. В качестве минорной линии присутствует у всех народов Средиземноморья, Ближнего Востока и Передней Азии. Ввиду своей редкости, на гаплокарте Азии включена в категорию «прочие». Согласно полевым исследованиям, не включенным в гаплокарту, является одной из основных у некоторых народов Эфиопии. Найдена также в ископаемой ДНК из Германии времен культуры линейно-ленточной керамики (7500-6800 лет назад), что косвенно говорит о раннем ее распространении в ходе миграций эпохи раннего неолита. Какие-либо детали истории гаплогруппы Т пока не удается реконструировать, и она остается одной из самых загадочных гаплогрупп современных людей.
В заключение, хотелось бы еще раз подчеркнуть, что представленный здесь обзор гаплогрупп не ставит своей целью дать исчерпывающую информацию по каждой из них. Его задача – очертить общую картину и отметить наиболее характерные черты. Дальнейшую информацию можно почерпнуть, изучая карты и специальную литературу, в том числе и в публикациях Академии ДНК-генеалогии. Приятного просмотра!
Игорь Рожанский,
кандидат химических наук
Перейти к авторской колонке

















