
На днях получил письмо от читателя: «Балановский обнародовал новую статью, она о датировках, скоростях, о ДНК-генеалогии и, как теперь обычно, Вам там уделено особое внимание. На мой взгляд, было бы интересно увидеть разбор этой статьи, она всё-таки о расчетах, чего раньше от попгенетиков не наблюдалось». Посмотрел. Опять типичная балановщина. Ключевые слова этого понятия – безграмотность, ложь, инсинуации. Собственно, балановщина – это и есть инсинуации вкупе с безграмотностью.

Фото: antropogenez.ru
Напомню определение понятия «инсинуация». Инсинуация – злостный вымысел, подстрекательство, преднамеренное сообщение ложных отрицательных сведений (измышление, клевета), выставление мыслей и поступков своего противника в таком свете, который может ему повредить и имеющее целью опорочить, порой подаваемое намёком (т.е. не прямо, а косвенным указанием на факты и обстоятельства). Цель инсинуации — подорвать в слушателях и/или читателях доверие к объекту инсинуаций, следовательно, к его доводам или поведению. Недобросовестность при инсинуации заключается в том, что инсинуирующий, зная действительное положение дел, по какой-то причине с умыслом создает у инсинуируемого или у окружающих ложное отрицательное представления о нём. При этом используются сложные подменные хитросплетения. В отличие от мистификации, инсинуация – это всегда акт в устном и/или письменном виде, и намерение носит не характер розыгрыша противника, а стремление его опорочить в глазах окружающих людей, поэтому в первую очередь и адресована им. Исполняется она в несколько этапов. Инсинуация всегда направлена против конкретного определенного человека, который представляется для инсинуирующего как препятствие к реализации своих целей и получению преимущества. При инсинуации применяются обороты речи в мягкой, внушающей форме, незаметно вкрадывающиеся в умы слушателей, для избежания их неудовольствия (из справочников и словарей).
«Новая статья» Балановского называется «Дискуссии о датировках», с подзаголовком «из книги О.П. Балановского «Генофонд Европы». Никакой дискуссии там, конечно, нет. Для дискуссии надо знать тему. Балановский о ней имеет представление крайне слабое, и поэтому это не «дискуссии», а примитивное изложение «по понятиям» некоего середнячка неких устоявшихся представлений некоего подобия науки под названием «популяционная генетика». «Подобия науки» – потому что научной школы в популяционной генетике нет. Поэтому на протяжении последних полутора-двух десятков лет попгенетики генерируют откровенный мусор в своих изданиях, о которых с придыханием говорят, что они «индексированные», «ваковские» и прочие. Это уже показывает уровень их требований к своей «науке». Я десятки лет работал в лучших научных школах СССР и США, и никогда не слышал, чтобы лидеры и научные сотрудники этих школ произносили слова типа «индексированное» или «ваковское» издание. Надо публиковать свои работы, и надо, чтобы другие их читали и использовали в последующих исследованиях. Надо входить в информационные научные потоки. А там мир сам разберется, что было важным, а что не оказало влияния на развитие науки. «Ваковское издание» – это сугубо бюрократическое изобретение.
 Переформат.ру пока находит силы держаться, но вопрос выживания сайта по-прежнему стоит очень остро. Чтобы продолжить «общественно-научное переформатирование» мы решили запустить проект на «Планете». Первые успехи были заслугой сравнительно небольшого круга наших читателей и сторонников, за что им сердечное спасибо (и презенты). Поэтому вопрос снова подвис. Даже от минимальной суммы, необходимой для продолжения Переформата, мы пока далеки. Переформат.ру: будет ли продолжение?.. Переформат.ру пока находит силы держаться, но вопрос выживания сайта по-прежнему стоит очень остро. Чтобы продолжить «общественно-научное переформатирование» мы решили запустить проект на «Планете». Первые успехи были заслугой сравнительно небольшого круга наших читателей и сторонников, за что им сердечное спасибо (и презенты). Поэтому вопрос снова подвис. Даже от минимальной суммы, необходимой для продолжения Переформата, мы пока далеки. Переформат.ру: будет ли продолжение?.. |
Вот и посмотрим на «дискуссии о датировках» Балановского, что там важно (или ничего?), что безграмотность, что инсинуации. Я не раз высказывал сожаление в публикациях, что попгенетики избегают обсуждений скоростей мутаций, потому что ничего в них не смыслят. Они ограничиваются общими словами, всегда негативными, никогда не дают конкретных примеров расчетов, а то, что в научных изданиях – у них всегда неверно, некорректно, неправильно. Впрочем, то же самое и в «дискуссии о датировках» Балановского – ни одного конкретного примера, опять общие слова.
Но это не просто так, не случайность. Балановскому очень хочется выглядеть «академичным», а в его системе координат «академичность» – это никогда ни на что не давать прямого ответа. Его «академичность» – это искажения, виляния, полуправда, но непременно с негативным исходом в отношении правды. Читать его измышления, честно говоря, противно, они производят отталкивающее впечатление. Возможно, потому, что они неряшливы, Балановский никогда не докапывается до первоисточников, и, не докопавшись, начинает искажать свою полуправду по своему усмотрению. Это – не академичность, это помесь безграмотности с обманом.
Прежде чем начинать систематическую порку Балановского, возьмем всего один пример из его «дискуссий» как иллюстрацию того, что сказано в предыдущих двух абзацах. Пример показательный. Он – о логарифмическом методе расчетов хронологии по гаплотипам, попросту говоря – расчет времени, когда жил общий предок рассматриваемой выборки гаплотипов. Этот метод с давних пор не дает покоя Балановскому. Причина проста – метод новый для тех, кто работают с ДНК, метод простой и эффективный. Но Балановский как середнячок, который сам ничего нового никогда не изобретал и не придумывал, ненавидит все новое и оригинальное. Для него все новое – раздражительный фактор. Казалось бы, если метод простой и эффективный – возьми на вооружение; если по каким-то причинам (например, по непониманию) не нравится – проходи мимо, не принимай на вооружение. Но нет, Балановскому непременно нужна инсинуация, дискредитация, нужно «разрушить», как ему представляется. Какая уж там «академичность»…
Кто не знает, что такое логарифмический метод – вот простой пример, который я опубликовал для пояснения метода в далеком 2008 году, в первом (июньском) выпуске Вестника Российской Академии ДНК-генеалогии, в статье под названием «Основные положения ДНК-генеалогии» на английском языке, и во втором (июльском) выпуске, уже на русском языке. Чтобы не загромождать читателя числами и расчетами, полезными больше для специалистов, вынесем это во врезку на странице. Кого детали не интересуют, можно пропустить, не читая. Вывод прост – логарифмический метод (в котором мутации не считают, считают только гаплотипы) дает верные и воспроизводимые результаты, совпадающие с линейным методом (в котором мутации считают) в пределах погрешности расчетов.
Еще в 2008 году я рассмотрел 68 гаплотипов шотландского семейства МакДоналдов гаплогруппы R1a, общий предок которых согласно документальным генеалогиям умер в 1386 году. Среди них было 53 одинаковых гаплотипа в 6-маркерном формате (а в остальных 15 гаплотипах суммарно было 17 мутаций от этих одинаковых гаплотипов), и 42 одинаковых гаплотипа в 12-маркерном формате (в остальных 26 гаплотипах было 44 мутации). В то время, семь лет назад, константа скорости мутаций для 6-маркерных гаплотипов была оценена как 0.0096 мутаций на гаплотип за условное поколение в 25 лет, а для 12-маркерных гаплотипов – как 0.024 мутаций на гаплотип за 25 лет. Логарифмическая формула дала для 6-маркерных гаплотипов время, когда жил общий предок [ln(68/53)]/0.0096 = 26±4 условных поколений, то есть 650±100 лет назад, и для 12-маркерных гаплотипов [ln(68/42)]/0.024 = 20±4 условных поколений, то есть 500±100 лет назад. Как мы видим, полученные величины находятся в пределах погрешности расчетов, неважно, 6- или 12-маркерные гаплотипы.
Более того, если применить линейный метод расчета, по мутациям в гаплотипах, то получим для 6-маркерных гаплотипов 17/68/0.0096 = 26±7 условных поколений (650±175 лет) до общего предка МакДоналдов, и для 12-маркерных гаплотипов 44/68/0.024 = 27±5 условных поколений (675±125 лет) до общего предка. Мы видим, что все эти величины близки друг к другу (формально – одинаковы в пределах погрешности расчетов), и они соответствуют документальной генеалогии семейства МакДоналдов гаплогруппы R1a, согласно которой их общий предок жил между 620 и 680 лет назад.
С тех пор немного подправились константы скоростей мутаций для 6- и 12-маркерных гаплотипов, после сравнительного изучения десятков и сотен серий гаплотипов, но метод, естественно, остался тем же, как и выводы из расчетов по гаплотипам МакДоналдов. Напомним, что метод был введен в июне 2008 года.
В октябре того же года в Вестнике № 4 была опубликована совместная статья (Адамов Д.С., Клёсов А.А. «Теоретическая и практическая оценка возвратных мутаций в гаплотипах Y-хромосомы») и следующей в том же выпуске была статья Д.С. Адамова «Расчет возраста популяции якутов, принадлежащих к гаплогруппе N1c1». В первой логарифмический метод не упоминался, только линейный, со ссылкой на мою статью в Вестнике № 2 (на русском языке). Во второй статье Д. Адамов использовал логарифмический метод, со словами «Используемый метод впервые был описан в работах А.А. Клёсова по основам ДНК-генеалогии», ссылка опять на Вестник № 2. После приведения формулы логарифмического метода, Д. Адамов сопровождает ее словами «Эта формула была впервые получена А.А. Клёсовым из уравнения кинетики первого порядка», ссылка опять на Вестник № 2.
Смотрим теперь на «новую статью» Балановского. Но сначала отметим ее стиль, который пронизывает всю статью. Это – ее неправдивость. Акценты «дискуссии» полностью смещены. Говоря о печально известной «популяционной скорости Животовского», ни слова о том, какое множество статей, в которых ее использовали попгенетики, пришли к неверным расчетам, выводам, интерпретациям, в том числе и статьи Балановских и их последователей. Везде, где Балановский заводит речь о ДНК-генеалогии, его стиль становится негативным, все достижения принижаются; везде, где речь идет о сомнительных методах попгенетики и тех, на которые попгенетика претендует, их оценка завышается. Это что, «академичный» подход? Нет, это просто ловкость рук. Но это хорошо для данного разбора «дискуссии» Балановского, материал для порки отличный. Во всяком случае, для образовательных целей.
Так вот, о логарифмическом методе. Что пишет Балановский? Фраза № 1 – «этот метод используется только в генетической генеалогии и в ДНК-генеалогии». Какая «генетическая генеалогия»? Кто в этом суррогате использует логарифмический метод? Где хотя бы один пример из «генетической генеалогии»? Нет таких. Вывод – Балановский не знает то, о чем пишет.
Фраза № 2 – «Этот метод… не был опубликован в признанных научных журналах». А какие «признанные» – определяет, конечно, сам Балановский. Статья, в которой логарифмический метод был опубликован еще в 2009 году, это статья в журнале Human Genetics, одном из ведущих журналов мира по генетике человека. Цитирую, в каком виде он там изложен:
Since all 98 haplotypes contain 16 base (identical to each other) 22-marker haplotypes (see above), it gives – employing the logarithmic method (see the last section of this paper) –ln(98/16)/0.047 = 39 generations (without a correction) or 41 generations (with the correction), i.e., 1,025 ybp to a common ancestor for Cohanim J1e* haplotypes. Since the logarithmic and the «linear» methods give the same dating to the common ancestor, it means that there was indeed just one common ancestor for the whole series of 98- of 22-marker haplotypes (see the last section), who lived about 1,075±130 ybp.
Перевод: «Поскольку все 98 гаплотипов содержат 16 базовых (идентичных друг другу) 22-маркерных гаплотипов, это дает – применяя логарифмический метод (см. последний раздел в этой статье) [ln(98/16)]/0.047 = 39 поколений (без поправки), или 41 поколение (с поправкой), то есть 1025 лет до общего предка для гаплотипов коэнов субклада J1e*. Поскольку логарифмический и «линейный» методы дали одинаковую датировку общего предка, это означает, что этот общий предок действительно существовал для всей серии из 98 гаплотипов в 22-маркерном формате (см. последний раздел), и он жил примерно 1075±130 лет назад».
С тех пор логарифмический метод был опубликован во многих статьях, и не прерогатива Балановского решать, «признанные» или «не признанные» там журналы. Например, статья, в которой был опубликован логарифмический метод, и в названии которой стоит «ДНК-генеалогия», была процитирована 34 раза

и еще одна статья – процитировано 29 раз,

Заметим, что целая обойма статей некой Е. Балановской до этого уровня цитируемости не дотягивает:

Ну так не затруднит еще раз повторить, что там про «признанные журналы»?
Поехали дальше. Фраза № 3 Балановского: «…я затрудняюсь привести точную ссылку – но среди пользователей метода общепринято считать, что он был разработан Дмитрием Адамовым и Анатолием Клёсовым». Здесь сразу три передергивания и искажения – (1) что он «затрудняется» привести точную сылку, хотя она доступна и выложена на сайтах, в том числе на сайте Академии ДНК-генеалогии, (2) что пишет «среди пользователей (логарифмического) метода», не поясняя, что это за такие таинственные «пользователи», которые что-то «общепринято считают» и, видимо, ставят об этом в известность Балановского; так вот, таких «пользователей» Балановский сам придумал, чтобы на них свалить свою безграмотность, и (3) про «общепринято считать» что метод был разработан… и так далее. Д.С. Адамов – грамотный математик, но логарифмический метод он не разрабатывал, см. выше. Он его использовал и ссылался при этом на мою работу, опубликованную ранее.
Думаю, любой уже понял, что Балановский литературу не читает, не знает, но вдохновенно импровизирует, передергивая и искажая. Это что, «академический подход»? Нет ничего дальше от него. Балановский – безграмотный ремесленник, не более того. Надо сказать, что дело вовсе не в авторстве метода, я за авторством не гонюсь, и уж тем более в логарифмическом методе, который я описывал еще в учебнике по химической и ферментативной кинетике для вузов в далеком 1976 году. Дело в том, что Балановский использует любой повод, чтобы исказить состояние дел. Это – не академический ученый, и рано или поздно его из науки погонят. На пользу и оздоровление самой науки.
Далее, Балановский пишет, опять про логарифмический метод – «Этот метод требует только знания того, какой гаплотип является исходным». Да ничего подобного! Когда среди 98 гаплотипов коэнов 16 были идентичны друг другу, то никакого «требует знания исходного гаплотипа» там нет. Ясно, что эти 16 – предковые гаплотипы, или базовые, что в данном случае одно и то же. Аналогично, когда среди 68 гаплотипов МакДоналдов 53 или 42 гаплотипа (в разном формате гаплотипов) были идентичны друг другу, то это тоже не требовало «знания исходного гаплотипа». Напротив, логарифмический подход это знание дает. Балановский не имеет об этом никакого понятия, поскольку сам этот метод никогда не применял. Потому и излагает «по своим понятиям», а не по науке, путая причину со следствием.
Чтобы закончить с логарифмическим методом, которому Балановский посвятил изрядную часть своей «новой статьи», пройдемся по другим свидетельствам его безграмотности. В своем страстном желании принизить значимость и удобство метода, Балановский начинает неумело и неквалифицированно «сравнивать» логарифмический метод с другими, о которых он тоже имеет неважное представление. Балановский пишет, что «таким же достоинством обладает и метод ASD». Опять, ничего подобного. Опять сказывается полное отсутствие практической работы, потому и получается ерунда. Заметим, что ни одного примера Балановский не приводит, по простой причине – он просто не владеет материалом. Только подумайте, с кем он надумал тягаться.
Так вот, простой пример – имеем 3466 гаплотипов в 111-маркерном формате, общее число аллелей 384 тысячи 726. Вопрос – сколько времени нужно, чтобы вручную найти все мутации от базового гаплотипа, и вычислить время до общего предка всей серии гаплотипов? И частный вопрос – сколько времени для этого понадобится Балановскому? Ответ – вопрос некорректный, он с этой задачей вообще не справится. Не тот уровень. Не та квалификация. Вопрос – сколько времени для решения задачи об общем предке понадобится мне? Ответ – c использованием логарифмического метода – несколько минут. Для этого, используя функцию «сортирование» в Excel, нахожу, что в 12-маркерном формате среди 3466 гаплотипов имеются 249 идентичных друг другу, то есть базовых, или предковых, гаплотипов. Получаем [ln(3466/249)]/0.02 = 132 → 152 условных поколений, то есть 3800±450 лет до общего предка. Это – первое преимущество логарифмического метода, а именно скорость и простота вычислений. Второе преимущество – возможность сопоставить результаты расчетов с линейным методом. Используя калькулятор Килина-Клёсова, получаем за секунду, что расстояние до общего предка всех 3466 гаплотипов в 111-маркерном формате равно 3810±381 лет, а в 67-маркерном формате 3841±384 лет (результаты расчетов умышленно не округляли). Разницы между логарифмическим и линейным (то есть по мутациям) методами расчета практически нет, на уровне сотых долей процента, хотя погрешность расчетов там умышленно завышена. Таким образом, второе преимущество, указанное выше, это возможность перекрестной проверки логарифмического и линейного методов. Если результаты совпали – значит, действительно выборка правильная, и формально вся серия из 3466 гаплотипов имеет одного общего предка, который жил 3800±400 лет назад.
Кстати, в качестве дополнительного «бонуса» калькулятор сообщает, что во всех 3466 гаплотипах число мутаций от базового гаплотипа равно 92120 в 111-маркерном формате, и 56200 в 67-маркерном формате.
Ну и какая цена сотрясению воздуха от Балановского – «таким же достоинством обладает и метод ASD». Это каким «таким же достоинством»? (ASD – это англоязычное сокращение от «усредненное квадратичное различие».) Не ответит Балановский, потому что знаний нет. Подобную же ерунду он пишет и далее – «ведь стоит в выборке случайно оказаться не двум, а трем или одному образцам с исходным гаплотипом, и его доля изменится в полтора-два раза, а значит, кардинально изменится и оценка возраста».
Откуда он взял «двум, трем или одному образцам»? Кто применяет в таких случаях логарифмический метод? Выше даны примеры для 53, 42, 16, 249 базовых гаплотипов в выборке, вот когда нужно применять логарифмический метод. Для двух «образцов» погрешность составит ±71%, для трех ±59%. Кому нужны такие расчеты, кроме попгенетиков?
Когда квалификация низкая, любой вопрос доводят до абсурда. Это напомнило мне давнюю лекцию по телевидению, когда лектор со знающим видом сообщал, что при езде на автомобиле на низкой передаче расход топлива выше, и привел пример, что если двигаться по шоссе на первой передаче со скоростью 60 км/час, то расход топлива будет большой. Вот Балановский – тот же по стилю лектор. Он заключает раздел о логарифмическом методе – «расчет по доле исходного гаплотипа не оптимален ни в каком случае». «По доле исходного гаплотипа» – это просто чтобы название «логарифмический метод» не использовать, пепел стучит в сердце Балановскому, когда речь о новых, прогрессивных подходах. В итоге он заговаривается, поскольку очень хочется принизить значимость, ну просто очень… Самое смешное, что об «оптимальных методах расчетов» рассуждает Балановский, ошибки у которого в расчетах датировок систематически равны как минимум 250-350%.
Я столько внимания уделил малограмотной писанине Балановского о логарифмическом методе, поскольку в этой «критике» отражается вся убогость познаний Балановского, и его манера вилять при рассмотрении любого вопроса, буквально как уж на вилах. Остальное в том же духе, но придется рассмотреть, хоть и противно.
Опустим пустую говорильню, которой заполнена «дискуссия» Балановского как минимум на три четверти. Примеров, как я уже упоминал, у него ни одного нет. Это – явный признак той самой пустой говорильни. Опять про «молекулярные часы», «где время отсчитывается происходящими мутациями». Добро бы он это сам придумал, так нет, просто обезьянничает. Я ему уже объяснял, что часы никому не нужны, если они время неправильное показывают. А чтобы «молекулярные часы» показывали правильное время, нужны выверенные константы скоростей мутаций. Вот этого у Балановского нет ни в статьях, ни в этой «дискуссии». Потому и пустая говорильня. Даже формулы для расчетов хронологии по мутациям бесполезны, если константы не определены и не выверены. В этом и есть принципиальный вклад ДНК-генеалогии – сопряжение расчетного аппарата с калиброванными и перекрестно проверенными константами скоростей мутаций. Об этом у Балановского – ни слова.
А что у него тогда вообще есть? Да ничего, та же пустая говорильня. «Если скорость мутаций постоянна, то часы будут работать», – изрекает он. Опять, кому такая «работа» нужна? Неверные часы тоже работают, отставая или торопясь на сотни минут, но кому такие нужны? Не «скорость мутаций постоянна», это само собой разумеется, и это в ДНК-генеалогии детально показано, что константы скоростей мутаций одинаковы в разных гаплогруппах и на любых реальных временах вплоть до миллионов лет назад, а то, что скорость мутаций должна быть выражена в конкретных величинах, и для индивидуальных констант скоростей для каждого из 111 маркеров, и для «кумулятивных» констант скоростей мутаций для гаплотипов любого заданного формата. Вот что нужно иметь для правильных расчетов. У Балановского и об этом ни слова. Как можно вообще без этого рассуждать о хронологических расчетах – уму непостижимо. Потому и у Балановского пустота – и информационная, и интеллектуальная. И в этой «новой статье», и вообще, по жизни, так сказать. Одно с другим связано.
Один за другим идут выхолощенные пассажи, типа «выявление и датировка времени их («кластеров») происхождения стали важным аспектом многих современных исследований Y-хромосомы». И вот так – вся статья. Ничего конкретного. Забавно, что первой ссылкой на пассаж выше идет ссылка на Л. Животовского, его печально известную работу 2004 года, в которой была введена «популяционная скорость», приводящая к завышениям датировок на 300% (!) (как соглашается Балановский, приводя это число). Уже больше десяти лет попгенетика с использованием этой «скорости» генерирует мусор в академической печати, что фактически похоронило попгенетику как науку. А Балановский не обучаем, опять приводит эту ссылку как основную, первую в серии. Сами ссылки, кроме номеров, в статье Балановского не приведены, но к неряшливости Балановского и попгенетиков мы уже привыкли. Что ни делают, всё тяп-ляп.
Повторяя зады попгенетики, Балановский утверждает, что «для STR гаплотипов Y-хромосомы… были предложены две скорости: “эволюционная” и “генеалогическая”». Опять неверно, нет «эволюционной скорости», это была грубая (и глупая) ошибка. Нет и «генеалогической скорости». Они разные для всех маркеров и гаплотипов. Для 6-маркерных гаплотипов скорость мутации равна в среднем 0.00123 мутаций на маркер на условное поколение, в то время как индивидуальные скорости мутаций для каждого из шести маркеров разные, и для DYS393 она равна 0.00059 мутаций на маркер за 25 лет (условное поколение), для DYS390 равна 0.00220 (в той же размерности), для DYD19 равна 0.00179, для DYS391 равна 0.00220, для DYS388 равна 0.00022, для DYS392 равна 0.00040. Какие там «генеалогические» скорости? Нет таких. Это каменный век попгенетики, который, впрочем, продолжается. Потому что нет у них научной школы.
Столь же примитивно описывается и метод расчета датировок с помощью ASD. В исполнении Балановского – это вообще верх бессмыслицы. Как пишет Балановский – для этого метода «Проводится расчет отдельно для каждого STR-маркера, а затем результаты по всем маркерам усредняются». Иначе говоря, берется некая смесь гаплотипов, невесть откуда появившихся, ничего о них знать не нужно, все сваливается в кучу, усредняется вдоль и поперек, и вуаля – получаем время до «общего предка» всей этой кучи, в которой этих предков – свой для каждой подкучи. Так «метод Животовского» и работал. Все усредняли и делили на «популяционную скорость» 0.00069 мутаций на маркер на 25 лет. Тоже был ASD. И получали завышения датировок в два-четыре раза, но могло быть в любое количество раз. Так в чем разница? Что, уже другую константу брать нужно, не «скорость Животовского»? Тогда какую? Нет, не даёт Балановский ответа. Не может он выйти за пределы «метода Животовского».
А на самом деле нельзя так делать. Надо решить две отдельных и независимых задачи. Первая – разделить эту исходную смесь гаплотипов на отдельные ветви, или «кластеры», у каждого из которых есть один, свой общий предок. Расчет надо вести для каждой ветви отдельно. Вторая – применить конкретную константу скорости мутаций для конкретных гаплотипов, из которых состоит каждая отдельная ветвь. Балановский же делает все чохом, без разделения на ветви, а какую константу скорости мутаций применять – не знает. Потому что он с этим не работает. Но беcтолковые «дискуссии» ведет, как ни поразительно. Это что, «академический подход»? Нет, это нахрапистая безграмотность.
Показательно, что Балановский тоже не имеет понятия в этом вопросе, хотя я это объяснял на протяжении последних лет. Но Балановский не читатель, он писатель. Он начинает раздел своей «дискуссии» со слов «для определения скорости мутирования… возможны два подхода: прямой подсчет и калибровка». Это – поразительная неосведомленность в элементарных вещах. Не будем повторять, что нет «скорости мутирования», есть константа скорости мутирования, потому что «скорость мутирования» без нормирования на что-то определенное и превращения таким образом в стандартный, научный показатель, никого не интересует. Опять, Балановский путается в простых понятиях потому, что не приводит конкретные примеры, а конкретные примеры он не приводит потому, что путается в элементарных понятиях. Круг замыкается.
Приведем несколько примеров расчетов, опять во врезке, чтобы не загружать неспециалистов деталями. Вывод первый – константа скорости мутации (для определенного маркера или определенного гаплотипа) на то и константа, что она верна для гаплотипов различных гаплогрупп. Вывод второй – Балановский не разбирается в том, что берется «рассматривать».
Пример 1 – в 100 гаплотипах в 25-маркерном формате за тысячу лет произошло 184 мутации. Где здесь «скорость мутирования»? 184 мутации за тысячу лет? Кого это интересует, если не проведено нормирование на сто гаплотипов, причем в 25-маркерном формате? А если оно проведено, то получаем 184/100/k = 1000 лет, то есть 40 условных поколений (по 25 лет). Здесь k – это константа скорости мутации в расчете на 25-маркерный гаплотип. Если рассчитывать ее на поколение, получаем, что k = 0.046 мутаций на 25-маркерный гаплотип на условное поколение. Это – именно константа скорости мутации, потому что она равна 0.046 при любом числе гаплотипов в серии и при любом числе мутаций в гаплотипах. Более того, та же константа скорости мутации определяет число базовых гаплотипов в выборке, для чего не надо и считать число мутаций, для этого есть тот самый логарифмический метод, который мы рассмотрели выше, и в чем Балановский тоже не разбирается.
Пример 2 – в серии из 661 гаплотипов гаплогруппы J1 в первых 25 маркерах имеется 3708 мутаций от базового гаплотипа, что дает «возраст» общего предка серии 3708/661/0.046 = 122 → 137 условных поколений, или 3425±350 лет (надо сразу сказать, что там совокупность разных ветвей, поэтому «общий предок» фантомный, как показывает логарифмический метод. Если отделить молодые ветви, то общий предок более старых ветвей жил 4000±450 лет назад, что, впрочем, в пределах погрешности с расчетом выше). Расчет по 111-маркерным гаплотипам дал 3245±330 лет, по 67-маркерным гаплотипам 3355±340 лет. Как видно, различия между этими тремя величинами попарно составляют от 2% до 6%, так что указанная погрешность явно завышена. Вот что такое константа скорости мутации, в данном случае 0.046 для 25-маркерных гаплотипов, 0.12 для 67-маркерных гаплотипов, 0.198 для 111-маркерных гаплотипов, во всех случаях ее размерность выражена в числе мутаций на гаплотип за условное поколение в 25 лет.
Пример 3 – в серии из 968 гаплотипов гаплогруппы I1 в первых 25 маркерах имеется 5656 мутаций от базового гаплотипа, что дает «возраст» общего предка серии 5656/968/0.046 = 127 → 144 условных поколения, или 3600±360 лет. Расчет по 111-маркерным гаплотипам дал 3690±370 лет, по 67-маркерным гаплотипам 3620±360 лет. Различия между этими тремя величинами попарно составляют от 0.5 до 2.5%. Опять, вот что такое константа скорости мутации.
Пример 4 – в серии из 164 гаплотипов гаплогруппы Т в первых 25 маркерах имеется 1748 мутаций от базового гаплотипа, что дает «возраст» общего предка серии 1748/164/0.046 = 232 → 293 условных поколения, или 7325±750 лет. Расчет по 111-маркерным гаплотипам дал 7200±725 лет, по 67-маркерным гаплотипам 7340±740 лет. Различия между этими тремя величинами попарно составляют от 0.2% до 1.7%. Опять, вот что такое константа скорости мутации. А «скорость мутации», о чем пишет Балановский, не понимая, что пишет – это 3708 мутаций в первом случае, или 5656 мутаций во втором, или 1748 мутаций в третьем, но без перевода в константы скорости мутации это ни о чем не говорит.
Итог – тяжелый случай с Балановским, и он еще книги садится писать по вопросу, в котором не разбирается.
По Балановскому, «прямой подсчет состоит в сравнении генотипов родителей и их потомства». Уже неряшливо, а по сути – неверно. Кстати, Балановский так и не написал, «прямой подсчет» – чего? Ну да ладно, косноязычность – это его визитная карточка. Здесь дело хуже. Генотип — это совокупность генов данного организма, а Балановский говорит о «STR-маркерах Y-хромосомы». Это та же неряшливость, с которой Балановские постоянно говорят про «генофонд», адресуясь к мутациям в нерекомбинантных участках Y-хромосомы.
Эти термины «генотип», «генофонд» в исполнении Балановских – или вопиющая безграмотность, или совершенно расхлябанный жаргон. Они, Балановские, повсюду используют термин «генофонд», относя его к гаплогруппам, субкладам и гаплотипам. Они не понимают, или не знают, или пренебрегают точностью понятий, что генофонд – это совокупность генов. Даже Википедия это знает, цитирую – «Генофонд (также генный пул, пул генов — англ. «gene pool»)». Понятие генофонда сформулировал еще в 1928 году А.С. Серебровский, цитирую – «Совокупность всех генов данного вида… я назвал генофондом». Нет в гаплотипах и гаплогруппах генов, никакой это не генофонд.
Балановские же от статьи к статье, от высказывания к высказыванию называют снипы (!) «генофондом». Очередной пример, из десятков и сотен, в недавней статье в сборнике «Археология, этнография и антропология Евразии» (Том 43, № 2, 2015), статья называется (привожу только часть названия) «Структура генофонда по данным маркеров Y-хромосомы». В авторах – Е. Балановская. И дальше, первая фраза Абстракта «Изучен генофонд популяции по SNP-маркерам Y-хромосомы». Категорически неверно. Я уже не говорю, что здесь безграмотность еще и в другом – не пишут в Абстракте «изучено то-то», пишут, что именно обнаружили, нашли, какие выводы сделали. Не пишут в Абстракте «мы собирали грибы», пишут «мы нашли 8 кг грибов», если уж объяснять на пальцах. Но Балановская и эти элементарные вещи не знает, научная школа напрочь отсутствует.
Они, конечно, сейчас начнут воздевать руки, причитать, что наука движется вперед, и А.С. Серебровский им не указ, уже и не гены в попгенетике называют генами – я с ними спорить на этот счет не собираюсь по простой причине – попгенетика себя уже настолько дискредитировала, что этот «генофонд» в применении к снипам всего лишь яркая заплата на ветхом рубище попгенетики, так сказать. Но яркая, показательная.
Возвращаемся по сути к некорректной фразе Балановского – «для определения скорости мутирования… возможны два подхода: прямой подсчет и калибровка». На самом деле это неверно – нет «прямого подсчета» без калибровки. «Прямой подсчет» – это, видимо, подсчет числа мутаций в парах отец-сын, хотя и этого Балановский не говорит, не умееет прямо излагать. По Балановскому, «хотя мутации случаются редко, но при больших выборках можно обнаружить достаточное их количество». Поскольку примеров он никогда не дает, изъясняется косноязычно, вязко, мутно, то не поясняет, что такое «при больших выборках» и что такое «достаточное количество». А это – ключевые понятия, о которых Балановский понятия не имеет, извините за каламбур.
Придется опять привести конкретные примеры, раз он не может, не владеет материалом. То, к чему призывает Балановский, было описано в статье Ballantyne и др. (2010), в которой на 1300-1800 парах отец-сын (такой разброс – потому что для каждого маркера число пар отец-сын различалось) определяли, сколько в каждом маркере произошло мутаций между отцом и сыном, то есть буквально за одно поколение. Авторы ставили своей задачей определить число мутаций за поколение для всех 111 маркеров, которые составляют общую панель маркеров. Но сразу же возникла проблема, о которой Балановский не упоминает ни слова, но мы уже привыкли к его уклончивому и вязкому стилю. Из 111 маркеров в итоге не изучались 24 маркера, в 17 маркерах мутаций вообще не было, в 15 маркерах прошла всего одна мутация – то есть 56 маркеров из 111 оказались совершенно непригодными для количественного определения скоростей мутаций. А поскольку еще в 11 маркерах прошли всего две мутации, то почти две трети всех маркеров были непригодны для определения констант скоростей мутаций. Даже при одной сигма (доверительный интервал плюс-минус 68%) погрешность в определении констант скоростей мутаций составляет ±100% при одной мутации, и ±71% при двух мутациях. А попгенетики их используют, в том числе и те маркеры, в которых мутаций вообще не было, при этом умудряясь рассчитать «скорости мутаций» для тех маркеров! В результате, разумеется, опять мусор в академических публикациях.
Пример такой работы – исследование 2013 года (Forensic Science International: Genetics. 7, 568-572), в авторах которого Chris Tyler-Smith, один из ведущих популяционных генетиков мира, и журнал один из ведущих. Я немедленно написал критическую статью в тот же журнал, и началась типичная для попгенетиков ситуация. Полгода ответа от журнала вообще не было. Я написал напоминание. После этого пришла одна рецензия, совершенно уклончивая, суть которой состояла в том, что несправедливо критиковать исследование, в котором используются мутации, определенные по парам отец-сын, поскольку многие их применяют. Поэтому моя статья быть принята не может. Я написал ответ, выразив возмущение сроками рецензии – более полугода, а также тем, что рецензент всего один, и само замечание неквалифицированное.
Через месяц пришла еще одна рецензия, в которой опять предлагалось снять критику за использование «скоростей мутаций» по парам отец-сын, снять таблицу, в которой показано, что значительная часть маркеров, используемых в работе Tyler-Smith, основывается всего на нескольких мутациях в парах отец сын. Так, по данным Ballantyne, в маркере DYS448 мутаций вообще не было (в 1747 парах отец-сын), в DYS438 прошла всего одна мутация (в 1751 паре), маркере DYS643 две мутации (в 1773 паре), и так в шести маркерах из 21, используемых в работе, более четверти. Помимо того, в работе использовались печально известные «популяционные скорости Животовского», которые вообще завышали датировки в три раза. Я ответил, что ничего снимать не буду. После этого получил письмо уже от главного редактора с приложением еще одной рецензии. Суть ее была в том, что несправедливо критиковать именно эту статью, и особенно несправедливо по отношению к Tyler-Smith, поскольку то, о чем я пишу, характерно по отношению буквально ко всем статьям популяционных генетиков, и почему начинать именно с Tyler-Smith? Поэтому мне предлагалось вообще снять всю критику данной статьи, и написать общую статью по мутациям в гаплотипах. Я отказался, написав, что сначала пусть они публикуют эту критическую статью, а потом обсудим более общую статью. После этого в течение года редактор мне периодически напоминал, что они ждут общую статью, но о критической статье не упоминал. Но я уже не отвечал. Такое отношение к авторам мне не подходит. И после этого попгенетики еще мне высказывают претензии, что я не публикуюсь в журналах по популяционной генетике. Нет уж, меня ангажированные издания не устраивают.
Резюмирую то, чего не понял Балановский. «Прямой подсчет» числа мутаций по парам отец-сын не подходит по ряду причин. Одна из них продемонстрирована выше – в объемной работе Ballantyne и др. (2010) из 111 маркеров данных не было получено для 24 маркеров, а для 56 данные совершенно ненадежны. Какая константа скорости для маркера, когда за поколение в маркере прошла всего одна мутация? Как константу будем рассчитывать? А если прошло всего 2-3 мутации, как константу рассчитывать будем? При погрешности в 70-100%? Пробовали бросить монету три раза, и по результатам рассчитать вероятность выпадения орла или решки?
А попгенетики так и не поняли, что эти данные по парам отец-сын для расчетов вообще непригодны. Возьмем, например, опять те же данные Ballantyne, потому что они самые массовые в одном эксперименте. В маркере DYS393, весьма «медленном», у 1750 пар отец-сын прошли три мутации, а в маркере DYS390, намного более «быстром», у 1758 пар прошли две (!) мутации, то есть маркер оказался как бы более медленный, что есть нонсенс. Любой, кто работает с гаплотипами, знает, что в DYS390 проходит больше мутаций, чем в DYS393. Вот пример – среди 3466 гаплотипов субклада R1b-L21 в маркере DYS393 от общего предка прошло 232 мутации, а в маркере DYS390 – 1164 мутации, в пять раз больше. А по парам отец-сын у Ballantyne – наоборот, 3 и 2 мутации, соответственно. Можно на основании расчетов по таким данным на что-то надеяться? Нет, конечно, и понятно, почему – статистика не та. Мало для расчетов 1700 пар отец-сын, очень мало. А Балановский, не имея понятия, пишет – «при больших выборках», «достаточное количество». Так какие выборки нужны? Какое количество достаточно, о чем Балановский, конечно, не знает? Уровень не тот.
Отвечу. Если даже взять 100 тысяч пар отец-сын, то те маркеры, у которых константа скорости мутации меньше чем 0.0001 мутаций на маркер на поколение, все равно покажут плохую статистику, у них в среднем будет наблюдаться меньше, чем 10 мутаций на поколение для всех этих 100 тысяч пар. То есть погрешность определения константы скорости мутации для них в лучшем случае в среднем будет ±32%. Но это только для тех, у которых константа скорости 0.0001. А таких, с константами 0.0001 и меньше, в 111-маркерной панели девять маркеров, из них пять – в 67-маркерной панели (показана константа скорости на маркер за условное поколение в 25 лет):
DYS472 0.000008
DYS436 0.000040
DYS425 0.000042
DYS568 0.000050
DYS490 0.000070
DYS426 0.000090
DYS455 0.000100
DYS632 0.000100
DYS494 0.000100
Кстати, для всех этих девяти маркеров в серии из 1500-1800 пар отец-сын (Ballantyne, 2010) не наблюдалось ни одной мутации. То есть их данные опять неприменимы для расчетов.
А для DYS472 и 100 тысяч пар-отец-сын недостаточно и для наблюдения хотя бы одной мутации. Там маркер мутирует в среднем раз в 125 тысяч поколений, то есть раз на 125 тысяч пар отец-сын. В общем, меньше чем полумиллионом пар не обойтись, лучше – миллион пар отец-сын. Это, конечно, практически нереально.
Но и там без калибровки не обойтись, несмотря на неквалифицированное заявление Балановского – «прямой подсчет и калибровка». Как мы видим, в ряде случаев в парах отец-сын наблюдаются ошибки, как показано выше. Или надо без всякой проверки их в дело пускать? Нет, придется все равно калибровать по известным генеалогиям или историческим событиям. И еще проблема, которую Балановский опять затронул, как всегда, уклончиво, нечетко, мутно. В парах отец-сын константы получаются в расчете на поколение. А в исторических науках на поколение не считают, нужно пересчитывать в года. А как? Популяционные генетики эту проблему так и не решили, см. следующий раздел. Так что в этом отношении пары отец-сын практически бесполезны. Польза, конечно, есть, но не для расчетов. Польза «концептуальная», просто посмотреть, какой порядок скоростей мутаций, и убедиться, что он «разумный». Вот, пожалуй, и вся польза. Но попгенетики до этого не додумываются.
Балановский пишет, что «таких исследований было проведено немало… результаты всех этих работ совпали, что является важнейшим подтверждением корректности этой оценки частоты мутаций». Ничего подобного, везде наблюдается разнобой. Балановского всегда приходится ловить за руку. Выше уже показано, какая там «корректность». Хорошо, сравним данные тех же Ballantyne и др. (2010) и большой, сводный список (по разным авторам) Burgarella и др. (2010), взяв для примера первые несколько маркеров из первой панели. Опять, чтобы не загромождать читателя информацией, полезной больше для специалистов, вынесем это во врезку на странице. Кого детали не интересуют, можно пропустить, не читая. Для них просто сообщу, что «частоты мутаций» у разных авторов по парам отец-сын различаются в два раза и более, и для расчетов они неприменимы. Никаких «совпали» и «корректности» там нет и близко. Балановский опять вводит в заблуждение, или попросту лжет.
DYS393, у Ballantyne 3 мутации на 1750 пар отец-сын, что в дальнейшем будем обозначать как 3/1750, у Burgarella 13/12576, то есть во втором случае 1.8/1750 мутаций вместо 3, в той же пропорции на 1750 пар, разница 167%.
DYS390, 2/1758 и 30/14131, соответственно, то есть во втором случае 3.7/1758 мутаций вместо 2, в той же пропорции, разница 185%.
DYS19, 7/1756 и 32/14632, соответственно, то есть во втором случае 3.8 мутаций вместо 7, в той же пропорции, разница 182%.
DYS391, константы скоростей мутаций оценены в диапазоне 0.00126-0.00665 (Ballantyne) и 0.00198-0.00372 (Burgarella). Формально диапазоны перекрываются, но вести расчеты с таким широким диапазоном неопределенности нельзя. По нашим независимым данным (Вестник Академии ДНК-генеалогии, № 3, 2015) величина константы скорости мутации для DYS391 равна 0.00220 мутаций на маркер на 25 лет.
DYS385a,b – данных у Burgarella нет, у Ballantyne дается диапазон 0.000624-0.00506 для DYS385a, то есть 8-кратный диапазон неопределенности, и 0.00175-0.00809 для DYS385b, то есть диапазон неопределенности в 4.6 раз. По нашим независимым данным (ссылка выше) величина константы скорости мутации для DYS385a и DYS385b равна 0.00280 и 0.00360 мутаций на маркер на 25 лет, соответственно.
DYS426 – у Burgarella данных нет, у Ballantyne ноль мутаций на 1735 пар отец-сын. Тем не менее, каким-то загадочным приемом последние оценили величину константы (при полном отсутствии мутаций) в 0.000398 мутаций на маркер на поколение (обратите внимание на точность оценки при отсутствии мутаций!). По нашим независимым данным (ссылка выше) величина константы скорости мутации для DYS426 равна 0.00009 мутаций на маркер на 25 лет, соответственно. Иначе говоря, Ballantyne в своей оценке промахнулись на 440%.
DYS388, 0/1635 и 1/2394 мутаций, соответственно, но и в такой ситуации (то есть при практическом отсутствии мутаций) Ballantyne и Burgarella оценили величины констант скоростей мутаций как 0.000425 и 0.0004177 на поколение, соответственно (опять обратите внимание на точность оценки!). Никаких оснований для таких оценок просто нет. По нашим независимым данным (ссылка выше) величина константы скорости мутации для DYS388 равна 0.00022 мутаций на маркер на 25 лет. Иначе говоря, Ballantyne и Burgarella в своих оценках промахнулись в два раза.
DYS439, 6/1736 и 51/9313 мутаций, соответственно, то есть во втором случае 9.5 мутаций вместо 6, в той же пропорции, разница 158%.
DYS389-1, 9/1751 и 32/12651 мутаций, соответственно, то есть во втором случае 4.4 мутаций вместо 9, в той же пропорции, разница 205%.
DYS392, константы скоростей мутаций 0.00097 и 0.0004303, соответственно, то есть разница 225%.
Теперь посмотрим на результаты по парам отец-сын в исследованиях Gusmao (2005) и Sanchez-Diaz (2008), на которые ссылался Балановский в своей «дискуссии». В первом исследовании изучали 3026 пар отец-сын по 17 маркерам, то есть должны были бы определить мутации в 3026х17 = 51442 маркерах-переходах от отца к сыну. На самом деле данные представлены всего для 27029 маркеров-переходов, то есть почти половину маркеров не изучали. В итоге сообщили, что константа скорости мутации в среднем на маркер находится в диапазоне доверительности между 0.001501 и 0.002606 за поколение (неизвестной продолжительности), то есть простирающимся на 174%. Ну и как, кому нужны расчеты с такой неопределенностью? К тому же в расчете на поколение неизвестной продолжительности. Кстати, по нашим независимым данным (ссылка выше) средняя величина константы скорости мутации для 17-маркерных гаплотипов (Y-filer) равна 0.00215 мутаций на маркер за 25 лет.
Смотрим на второе исследование, в котором изучали 701 пару отец-сын, и нашли в 17-маркерных гаплотипах суммарно 26 мутаций. Уже ясно, какая там статистика. Да, так и есть. В Абстракт авторы вынесли, что для DYS438 константа скорости мутации равна 0.000425 на поколение (опять неизвестной продолжительности), а в работе Ballantyne для 1751 пар отец-сын та же константа скорости мутации равна 0.000956 на поколение. Разница – 225%. Ну ладно, этот маркер «медленный», хотя в 17-маркерную панель Y-Filer он входит. Но посмотрим на самый быстрый маркер этой панели, DYS458. На 701 паре отец-сын константа равна 0.00636, у Ballantyne на 1756 пар константа равна 0.00838, разница на 31%. И это – самая благоприятная статистика, «быстрый маркер». Как там у Балановского – «результаты совпали»? «Важнейшее подтверждение корректности»? Он на данные в статьях смотрел или нет?
Пожалуй, достаточно. Вывод – «частоты мутаций» по парам отец-сын практически невоспроизводимы, никаких расчетов с ними делать нельзя.
Любой, хоть немного понимающий в числах, оценит лживость заявления Балановского, что «результаты всех этих работ совпали, что является важнейшим подтверждением корректности этой оценки частоты мутаций». На самом деле ситуация ясна – Балановский на результаты и не смотрел, он из тех, кому результаты и не нужны. Он – политработник, так сказать. Если проще – работающий по некому уставу, где мысли и не нужны. Есть внутренние установки, которые надо выполнять. Нет ничего, более далекого от науки. Балановский – не ученый, он техник-лаборант некоего негативного идеологического фронта. Поэтому научные дискуссии с ним бесполезны.
Сюда же относятся его абсолютно ненаучные рассуждения о якобы «калибровке», проведенной Л. Животовским (2004) в отношении «популяционных скоростей». Опять Балановский заводит пластинку о якобы «калибровке» по гаплотипам маори и цыган, которую якобы проводил Животовский. Эта «калибровка» детально рассмотрена мной в ряде журнальных публикаций, и в книге «Происхождение славян», глава 16. Не было там никакой «калибровки». Была откровенная подгонка данных с серийным выбрасыванием неугодных гаплотипов, причем подгонка шла в несколько кругов, и в итоге все равно подогнать не удавалось, и нужное число просто декларировалось. Либо Балановский этого не читал, либо просто лжет. Я не знаю, что хуже.
И после этого Балановский «на голубом глазу» объявляет, что «к сожалению, других работ по калибровке скорости мутирования Y-STR маркеров не было проведено». Здравствуйте, приехали. А как же статья в «Advances in Anthropology» (Rozhanskii and Klyosov, 2011) c калибровками по тысячам гаплотипов из разных гаплогрупп? А как насчет калибровки еще в 2008 году по гаплотипам МакДоналдов, опубликованной в Вестнике Академии ДНК-генеалогии? В ДНК-генеалогии еще с 2008 года используются откалиброванные значения констант скоростей мутаций гаплотипов разных форматов, описанные в книгах «Происхождение человека» (2010), «Происхождение славян» (2013), «Занимательная ДНК-генеалогия» (2013), «Арийские народы на просторах Евразии» (2015), «Славяне, кавказцы, евреи с точки зрения ДНК-генеалогии» (2015), «Евреи и пуштуны Афганистана» (2015), «Экспертиза Велесовой книги» (том третий, 2015). Как это Балановский все пропустил? Ах, да, Балановский – не читатель, Балановский – писатель…
О калибровке констант скоростей мутаций Балановский тоже не имеет понятия. Взгляните на то, что он пишет: «Калибровка состоит в определении разнообразия гаплотипов, накопленного популяцией за время ее существования». Никакого отношения к калибровке «определение разнообразия» не имеет. Что такое «разнообразие гаплотипов» он тоже не поясняет. Поскольку он косноязычен и невразумителен, приходится за него догадываться, что он имеет в виду. Разнообразие – это, наверное, число мутаций в серии гаплотипов. Но как ни догадывайся, к калибровке никакое «разнообразие» не имеет отношения. Более того, выражение «накопленного популяцией за время ее существования» – это что, как ни косноязычность и невразумительность? Что, всей популяцией? Сотнями тысяч и миллионами человек? Это – наглядный пример, что Балановский не имеет никакого опыта работы с гаплотипами и мутациями. Не популяцией накопленного «за время ее существования», это вообще нонсенс, а речь должна идти о выборке.
Но не только это косноязычие делает тексты Балановского нонсенсом. У него процитированная фраза еще более бессмысленна. Вот простой пример – имеем два набора по три 17-маркерных гаплотипа, один набор «разнообразнее», другой «менее разнообразен», или, точнее, вообще не разнообразен:
13 24 14 10 11 14 12 12 13 28 17 15 19 12 16 12 23
13 23 14 11 11 15 13 13 13 29 18 16 19 12 15 12 25
13 23 14 13 11 14 12 14 13 30 17 15 19 13 16 12 23
13 24 14 11 11 14 12 13 13 30 17 15 19 12 16 12 23
13 24 14 11 11 14 12 13 13 30 17 15 19 12 16 12 23
13 24 14 11 11 14 12 13 13 30 17 15 19 12 16 12 23
Это, кстати, гаплотипы басков. Первая серия – три реальных гаплотипа, вторая – три базовых гаплотипа басков. Ну, определили «разнообразие» гаплотипов. Выразили их в числе мутаций. Дальше что? Где калибровка? Откуда она следует? Повторю опять безграмотную цитату из Балановского – «Калибровка состоит в определении разнообразия гаплотипов, накопленного популяцией за время ее существования». С таким определением попгенетика и стала паранаукой.
На самом деле у этого «определения» Балановского отсутствует ключевое положение – калибровка это не «определение разнообразия», это выражение выявленного «разнообразия», как правило, в числе мутаций от базового маркера или базового гаплотипа, в количестве лет до общего предка, которое находится независимо.
После проведения большой работы были определены константы скорости мутации для каждого из 111 маркеров, используемых в ДНК-генеалогии (Килин и Клёсов, Вестник Академии ДНК-генеалогии, № 3, 2015). Значения этих констант были выверены по многим документальным генеалогиям и другим независимым временным показателям, и, в частности, показано, что кумулятивная константа скорости для 17-маркерных гаплотипов (содержащих пару маркеров DYS385) равна 0.0365 мутаций на гаплотип на условное поколение в 25 лет. Вот что такое калибровка, а не «определение разнообразия». «Определения разнообразия» без введения временной компоненты ничего не стоит, ничего из него не получить.
Так вот, три верхних гаплотипа содержат 15 мутаций от базового гаплотипа. Статистика неважная, но для демонстрационных целей подходит. Получаем 15/3/0.0365 = 137 → 160 условных поколений, то есть примерно 4000 лет до общего предка этой выборки из трех гаплотипов басков. Для справки – 272 гаплотипа басков в 17-маркерном формате показывают 1478 мутаций от базового гаплотипа, что дает 1478/272/0.0365 = 149 → 177 условных поколений, или 4425±460 лет до общего предка. Но чтобы это получить, надо было получить откалиброванную константу скорости мутаций для 17-маркерных гаплотипов, а не определить «разнообразие». Итог – опять Балановский не понимает, что пишет.
Балановский, конечно, скажет, что после цитированной выше фразы у него стоит «для этого нужно изучить популяцию, для которой известно время ее основания». Но эта фраза тоже практически бессмысленна, потому что в попгенетике такие популяции не изучают, наоборот, «время основания» принимают совершенно безосновательно. Так поступал Л. Животовский (2004), который принял, что маори (полинезийцы) прибыли на острова 800 лет назад, и заложил это число в «калибровку». Но так делать категорически нельзя. Попгенетики, и Балановский с Животовским с ними, не понимают простого положения, что прибыть маори могли 800 лет назад, но это вовсе не означает, что это есть начало их ДНК-генеалогической линии. Если они прибыли группой, то «принесли» общего предка популяции с собой. Иначе в США гаплотипы группы R1b все имели бы общего предка в 1620 году. Далее, маори могли прибыть на острова Кука и Самоа 800 лет назад, но пройти бутылочное горлышко популяции, и общий предок их мог жить намного позже. Как можно закладывать в этой ситуации 800 лет назад как начало ДНК-генеалогической линии? Вот этот примитивизм мышления и интерпретации данных в попгенетике уже не удивляет. Паранаука.
Потому-то у Животовского и получилась ерунда при такой «калибровке». Расскажем как курьез, как у Животовского эта «калибровка» проходила. Были взяты 22 гаплотипа маори и 23 гаплотипа с островов Кука, в виде серии 10-маркерных гаплотипов у маори, 10 и 7-маркерные гаплотипов у островитян Кука, и 10-маркерных гаплотипов у четырех самоанцев, и постулировано, что полинезийцы прибыли на острова 32 поколения назад, при 25 годах на поколение, то есть 800 лет назад. Применяя квадратичный метод, авторы показали, что среднее число мутаций на маркер у маори и островитян Кука равно 0.00998, при этом самоанцев в расчет не включали, а маори и островитян считали по 7-маркерным гаплотипам. По 10-маркерным расчета (у маори, или маори и самоанцев вместе) не было, видимо, что-то не стыковалось. На основании этого авторы посчитали, что «скорость мутации» равна 0.00998/32 = 0.000312 на маркер на поколение. А надо было получить 0.00069 на маркер на поколение, просто задача была такая, это было «математически предсказано», заметим, что при совершенно искусственных условиях. Но 0.000312 – это вовсе не 0.00069, а в два раза медленнее.
Далее авторы заключили, что это, скорее всего, недооценка, и опять пустились в долгие и вязкие рассуждения, почему это может быть недооценкой. Потом решили ввести «веса» в исходные серии гаплотипов, что привело авторов к «корректировке» средней скорости мутаций в серии полинезийцев от 0.000312 до 0.000705 (!), то есть более чем в два раза. Более того, с учетом погрешности таких поправок величина средней скорости мутации стала равна 0.000705±0.000332, со стандартным отклонением по маркерам 0.00078, то есть выше, чем сама скорость мутации. На этом «калибровка» этой серии гаплотипов была завершена, и принято, что «скорость мутации» равна 0.00069, что и требовалось доказать. Это и есть «калибровка» у попгенетиков.
Подобный же постулат был принят и при «калибровке» с гаплотипами цыган Болгарии. Животовский с соавторами сообщили, что цыгане были замечены в Болгарии 700 лет назад, то есть 28 поколений назад, и это тоже было принято в качестве основы для калибровки. Как так можно? Но для попгенетиков все можно. Они сообщили, что анализировали 179 гаплотипов из 12 цыганских таборов в Болгарии. И вот как проходила «калибровка». Сначала сообщили, что один гаплотип (единственный из табора Даракчи) был исключен, потому что оказался нечетным (? – ААК), затем усреднили число мутаций по всем 8-маркерным гаплотипам, получили 0.01272 мутаций на маркер, разделили на постулированные 28 поколений, и получили 0.000454 как среднюю скорость мутаций на маркер. Отметим, что это опять заметно ниже, чем якобы «калиброванная» 0.00069.
Авторы опять стали обсуждать, что это должна быть «недооценка». Далее опять внесли «коррективы», а именно «веса» в исходные серии гаплотипов и сняли ряд гаплотипов цыган. Так, авторы решили, что один табор – «Музыканты» – очень «гетерогенный», потому что их гаплотипы отличаются на две аллели в шести гаплотипах из 19 от наиболее распространенного гаплотипа. Все 19 гаплотипов «Музыкантов» сняли со счета и опять ввели «веса» по размеру популяции (что совершенно нельзя делать в случае вполне однородной серии гаплотипов, какой серия гаплотипов цыган и была, никакой «генерогенности» там не было, «Музыкантов» нельзя было снимать, дерево гаплотипов и с ними было симметричным, но авторы деревьев не строили). Получили среднюю скорость мутации 0.000725±0.000187, и опять решили, что это и есть искомые 0.00069. Если это «калибровка», тогда непонятно, что такое не калибровка. На самом деле это откровенная подгонка.
И сейчас Балановский опять твердит о том же подходе, про «популяцию, для которой известно время ее основания», и – цитирую – «скорость мутирования STR-маркеров Y-хромосомы была определена в работе (Животовский, 2004) на двух примерах… популяции маори… в результате миграции полинезийцев не позднее 800 лет назад, и популяции цыган Болгарии… около 900-1000 лет назад». Но мы уже видели, что там была за калибровка. При этом Балановский пишет – «к сожалению, других работ по калибровке скорости мутирования Y-STR маркеров не было проведено, поэтому отсутствуют независимые подтверждения (как, впрочем, и опровержения) этой скорости» (то есть «скорости Животовского).
То есть Балановский делает вид, что не знает о серии работ по калибровкам констант скоростей мутаций в ДНК-генеалогии за последние 8 лет, и о существовании главы 16 в книге «Происхождение славян» (М., 2013), в которой все эти «калибровки» Животовского на многих страницах были детально разобраны и отвергнуты, данные по маори и цыганам пересчитаны, и показано, что никакой «скорости Животовского», она же «популяционная скорость», не существует. А Балановский без стеснения к ней опять возвращается. Разумеется, ни слова про огромный ущерб от этой «популяционной скорости», который фактически погубил популяционную генетику как науку, у Балановского нет. Как нет ни слова и про ошибочные статьи самих Балановских, которые активно использовали этот неверный «метод Животовского». И понятно, почему – как только он в этом сознается, встанет вопрос про отзыв многих статей Балановских и их последователей.
Нет, Балановский определенно необучаем. Последние 8 лет я объясняю, что продолжительность поколения автоматически входит в константу скорости мутации, потому что при делении числа мутаций на число гаплотипов в серии автоматически получается произведение kt, где k – константа скорости мутации, t – время, прошедшее от общего предка рассматриваемой серии гаплотипов (при условии, конечно, что предок был один, и это позволяет установить логарифмический метод в паре с линейным). Как я уже пояснял, время удобнее выражать в условном числе поколений, потому что в годах величина k будет иметь много нулей, что создает практические неудобства. Видно, что какую продолжительность поколения ни брать, константа скорости мутации сама подстроится, при известной величине kt, которая и определяется калибровкой. Если взять продолжительность поколения в 25 лет (потому она и «условная», потому что принимается как математическая величина, вместо «плавающей» абсолютной величины поколения в годах, которая зависит от культурных, религиозных, бытовых факторов, войн, эпидемий и проч., и бессмысленно ее пытаться определять в науке; ее постулируют правительственные и неправительственные огранизации, такие, как ООН, ЮНЕСКО и прочие, для целей скорее организационных). Величины кумулятивных констант скоростей мутаций, приведенные выше, как 0.02 мутаций на условное поколение (25 лет) для 12-маркерных гаплотипов, 0.0365 для 17-маркерных, 0.046 для 25-маркерных, 0.09 для 37-маркерных, 0.12 для 67-маркерных, 0.198 для 111-маркерных – они все откалиброваны для 25 лет на условное поколение. Если по какой-либо загадочной причине взять другую продолжительность поколения, то константы скорости перестроятся, адаптируются, и конечный результат получится точно таким же.
Приведу пример. Для 67-маркерных гаплотипов средняя константа скорости мутаций равна 0.12 на гаплотип на условное поколение в 25 лет. Поэтому при 120 мутаций на 50 гаплотипов общий предок этих 50 гаплотипов жил 120/50/0.12 = 20 условных поколений назад, то есть 20х25 = 500 лет назад. Если кому-то не нравится величина 25 лет на условное поколение (правда, причины такой нелюбви к 25 годам на поколение будут загадочными), и этот некто захочет использовать 30 лет на поколение, то нет проблем, просто константу скорости мутации придется пропорционально пересчитать, и она получится 0.12х30/25 = 0.144 мутаций на поколение в 30 лет. Тогда для 120 мутаций имеем 120/50/0.144 = 16.666 поколений по 30 лет, или те же 500 лет до общего предка. От чего ушли, к тому же и пришли.
А что пишет Балановский, несмотря на мои многолетние разъяснения? Ах, да, он же не читатель, он писатель. К тому же писатель необучаемый. Он пишет – «В данном исследовании во всех методах нам пришлось пользоваться двумя разными величинами длины поколения для двух мутационных скоростей. При использовании эволюционной скорости мутирования, длина поколения принималась равной 25 лет… (Животовский, 2004). При использовании генеалогической скорости мутирования, длина поколения принималась равной 30 годам…». Мало того, что он продолжает держаться за «эволюционную скорость» Животовского, что никакого смысла не имеет, он продолжает не понимать, что 30 лет на поколение по определению должно входить в «генеалогическую скорость», а для Балановского «генеалогическая скорость» – нечто застывшее, одинаковое для всех маркеров. Опять приходится повторить, что он так и не понял, что в гаплотипы разных форматов входят свои комбинации маркеров, и у каждого своя константа скорости мутаций, а значит, нет некой одной «генеалогической скорости», они все разные. Но у попгенетики свои представления, от науки далекие. И культивируют их, в частности, Балановские.
В своей «дискуссии» Балановский затронул и датировки по снипам. Надо сказать, что отпечаток примитивизма и непонимания сути вопроса в полной мере присущ и этой части статьи. Это становится ясным уже в самом начале изложения, когда Балановский пишет – «Метод расчета возраста гаплогрупп по полногеномным данным столь прозрачен, что обычно даже не обсуждается». Не обсуждается только среди таких же примитивов. На самом деле, в этом методе масса неясностей, подводных камней, недоговоренностей, технических проблем. Без сомнения, метод датировки по снипам – крупный шаг вперед, но он еще не готов для надежного применения. Балановский в силу своей ограниченности видит только один аспект этого метода – выявление средней скорости мутаций на нуклеотид в год, которая в последний год-два постепенно сходится к величине 0.8х10-9 на нуклеотид в год, как цитирует Балановский. И Балановский в силу той самой ограниченности, присущей типичному технику-лаборанту, считает, что дело сделано, метод создан. Ничего подобного, это только начало.
А дальше – две огромных неясности: сколько именно снип-мутаций ведет к тому узлу (нужному снипу), датировку которого нужно определить, и сколько лет приходится на снип-мутацию в каждом конкретном случае. С этим связана еще одна неясность – какой средний размер Y-хромосомы анализируется, и какое там распределение по размерам фрагментов. Последний фактор по сути углубляет и первую, и вторую неясность. А средняя скорость мутаций, 0.8х10-9 на нуклеотид в год, которую Балановский только и рассматривает, оказывается самым маловажным параметром, поскольку число нуклеотидов очень часто остается неопределенным – ну, и куда эту скорость привязать? Вот и получается погрешность расчетов по снипам зачастую на уровне ±30%-50%, часто и намного больше. А у Балановского – «столь прозрачен, что обычно даже не обсуждается». Остается только руками развести от такого примитивизма «мысли».
Поясню на нескольких примерах. Ниже приведена диаграмма Y-хромосомных линий для 77 носителей субклада R1a-Z280. Числа в нижней части диаграммы показывают число снипов от узла Z280 до настоящего времени для каждого из 77 человек. В идеале это число снипов должно быть у всех одинаково, но в реальности картина сложнее. Никакой «прозрачности, что даже не обсуждается», как представляется Балановскому с его ограниченными представлениями, там нет и близко. На самом деле это число снипов варьируется от 22 (один человек) до 45 (два человека). Медиана – 36 снипов (7 человек), максимальное число человек (10) имеют 37 снипов, средняя величина – 35 снипов. Как считать будем, если все так прозрачно? Куда будем вставлять среднюю скорость мутации 0.8х10-9 на нуклеотид в год, которую приводит Балановский?
Как он пишет, если секвенируют фрагмент Y-хромосомы размером в 10 миллионов нуклеотидов, то «одна мутация произойдет в среднем за 120 лет… значит, за тысячу лет… произойдет уже целых восемь мутаций, то есть по числу мутаций можно оценить возраст гаплогруппы с приемлемой статистической погрешностью». Балановский в присущей ему вязкой, неквалифицированной манере изложения не поясняет, что такое «приемлемая статистическая погрешность», так что придется нам опять за него это показать. Итак, если считать по средней величине, то снип Z280 образовался 35х120 = 4200 лет назад, если по медиане, то 4320 лет назад, если по максимальному представительству, то 4440 лет назад. Но это без разброса по числу снипов, и так считать неправильно, а на самом деле должно быть между 2640 и 5400 лет назад. Это «приемлемая статистическая погрешность», или как? И все ли там «так прозрачно, что даже не обсуждается»? Так и про «скорость Животовского» писали, а сейчас Балановский чешет голову, и говорит примерно: «Как же так? Разница-то в три раза»… У попгенетиков все так, «прозрачно».
Смотрим данные компании YFull, когда образовался снип Z280 по их подсчетам.
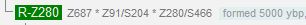
У них – 5000 лет назад. Как же так, если все так «прозрачно», что и обсуждать нечего? В чем причина? Да в том, что они берут, видимо, некое свое число снипов, одна снип-мутация у них принимается не за 120 лет, а за 144 года, и фрагмент Y-хромосомы они принимают не за 10 миллионов нуклеотидов, а за 8.47 миллионов, см. ниже. Нетрудно подсчитать, что они берут за основу примерно среднюю величину, около 35 снип-мутаций, хотя их там, как сообщено выше, между 22 и 45.
Так что у Балановского весьма примитивное представление о том, как ведутся расчеты датировок по снипам. Иллюстрация ниже по клику открывается в увеличенном виде в новом окне.
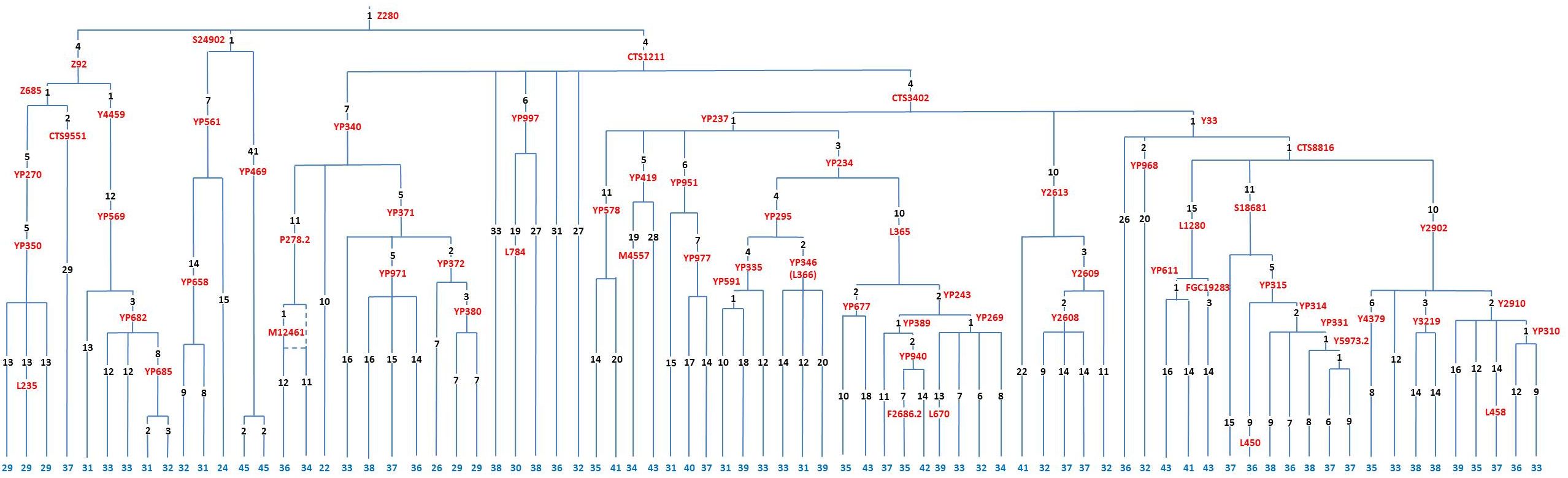
Но там еще менее «прозрачно», чем то, что мы сейчас показали. Смотрим на датировки образования гаплогрупп R1a и R1b по данным компании YFull.
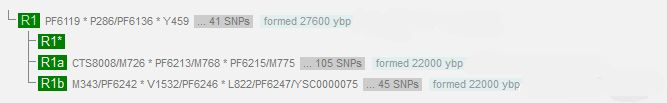
Видим, что к R1a ведут 105 снип, к R1b – 45 снипов. Сама подача данных совершенно невразумительна, поскольку при 144 годах на снип это дает 15120 и 6480 лет, соответственно. Похоже, что это расстояния от датировки гаплогруппы R1, на что намекает разница в (27600-22000) = 5600 лет, что в целом недалеко от гаплогруппы R1b. А гаплогруппа R1a, что с ее датировкой? 27600-15120 = 12480 лет? А в таблице стоит та же 22000 лет, что и у R1b. Ах, да, по Балановскому там «столь прозрачно, что обычно даже не обсуждается». Всем понятно, что Балановский во всём натягивает, передергивает, жульничает? Это – его уровень, его квалификация. Его визитная карточка.
Теперь – мой взгляд на состояние датировки по снипам, поскольку Балановского слушать нельзя, нигде и ни в чем. По данным разных исследователей, необратимые снип-мутации в Y-хромосоме, и, видимо, в ДНК в целом, происходят со средней скоростью между 0.5х10-9 и 1.0 х10-9 на нуклеотид в год. В последнее время прошла серия работ независимых исследователей, в которых средняя скорость снип-мутаций определена как 0.78-0.82х10-9 на нуклеотид в год. Наконец, исследователи в компании YFull, которые занимаются расчетами скоростей мутаций в Y-хромосоме (Адамов, Гурьянов, Каржавин, Таганкин, Урасин, 2015, «Константа скорости SNP мутаций Y-хромосомы по данным полного секвенирования»), принимают в своих расчетах величину скорости 0.8178×10-9 на нуклеотид за год, хотя такая точность определения совершенно нереальна. Пагубное влияние попгенетики. Поскольку вся Y-хромосома – примерно 58 миллионов нуклеотидов, и если скорость снип-мутации – 0.8178×10-9 на нуклеотид за год, то это составляет 0.8178х10-9х58х106 = 0.04743 на всю хромосому в год, или 1.185 на всю хромосому за 25 лет. Даже если скорость мутаций не такая точная, а, как свидетельствуют разные публикации, между 0.5 и 1.0х10-9, то снип-мутации происходят в среднем между 0.725 и 1.45 мутаций на всю Y-хромосому за 25 лет. То есть, как ни считать, мутации в Y-хромосоме происходят со средней скоростью примерно одна мутация за поколение.
Сформулируем по-другому – если скорость мутации брать от 0.5х10-9 до 1.0х10-9 на нуклеотид за год, то снип-мутация происходит раз в интервал времени между 34 и 17 лет. Мы уже видим, что если скорость мутации не откалибровать надежно, то точность метода датировки по снипам невелика. Помимо того, точность в реальных условиях еще более низкая, потому что почти никто не секвенирует полную Y-хромосому в поисках снипов, а секвенируют фрагменты, либо небольшие, в 300-500 тысяч нуклеотидов, либо в 10 миллионов нуклеотидов (как в методе BigY), либо, реже, в 30 миллионов нуклеотидов. Но когда пишут, что секвенируют фрагмент в 10 миллионов нуклеотидов, там фрагмент легко может оказаться между 8 и 11 миллионами, и это еще оптимистическое предположение.
Давайте посмотрим, к чему это приведет. Как отмечалось выше, компания YFull принимает величину константы скорости мутации 0.8178×10-9, в таком случае одна мутация во всей Y-хромосоме происходит в среднем раз в 1/0.04743 = 21.08 лет. Во фрагменте в 30 миллионов нуклеотидов – раз в 41 год. Во фрагменте в 10 миллионов нуклеотидов – раз в 122 лет. Если фрагменты малые, в 300 тысяч нуклеотидов, то раз в 4075 лет. Вот и выбирайте. YFull собщает, что средний размер фрагмента Y-хромосомы, по которому они ведут расчеты, составляет 8467165 нуклеотидов (такая точность тоже смешна, ну да ладно, это у попгенетиков неисправимо). Тогда одна мутация в таком фрагменте происходит в среднем раз в 144.41 лет (это опять сообщают YFull, опять с такой точностью). Когда умножают на число снипов, к этому прибавляют еще 60 лет. Как объясняют в YFull, 60 лет – это средний возраст клиента, который к ним обращается. Такая «точность» тоже вызывает усмешку.
Итак, если читатель не хочет разбираться в деталях, то на этом этапе можно резюмировать, что если исследователи сообщают, что анализируют (в поисках снипов) фрагменты Y-хромосомы в 10 миллионов нуклеотидов, то там на самом деле может быть восемь с половиной миллионов нуклеотидов, или меньше, или больше, и если при этом считать, что скорость снип-мутации составляет именно 0.8178×10-9 на нуклеотид в год, то при фрагменте в 8.5 миллионов нуклеотидов одна снип-мутация происходит раз в 144 года. На деле там, конечно, погрешность более чем солидная.
Но это еще не всё, и погрешность часто еще значительно больше. Дело в том, что снип-мутации происходят не через равные промежутки времени, а неупорядоченно. Если при этом рассматриваются цепочки снипов протяженностью десятки и сотни тысяч лет, то это аналогично рассмотрению ситуации с сотнями бросков монеты, и погрешности из-за неупорядоченности нивелируются. Но для относительно недавних снипов метод расчета очень неточный. Например, если снип образовался 750 лет назад, то в цепочке нисходящих снипов их всего 750/144 = 5 cнипов, и погрешность определения времени образования снипа, согласно формулам математической статистики, не меньше ±46% (с 68%-ной достоверностью) или ±92% (с 95%ной достоверностью). В итоге погрешности у большинства датировок по снипам в таблице значительно более высокие, чем у датировок по мутациям в гаплотипах, которые рассматривает ДНК-генеалогия.
Есть еще серьезная проблема – в расчетах компании YFull часто игнорируют фактическое количество снипов, и ставят ту датировку, которая им представлется более правильной, «по понятиям». При этом они всегда постулируют, что при разветвлении снипов они образовались точно в одно и то же время, хотя это события совершенно независимые. По аналогии, если у отца двое сыновей, то их возраст далеко не всегда одинаковый, один мог родиться, когда отцу было 18 лет, другой – когда отцу было 70 лет. Такой постулат, «о равенстве возраста сыновей» часто приводит к ситуациям, когда к одному снипу от вышестоящего ведет 5 мутаций (в среднем 720 лет), к другому 20 мутаций (в среднем 2880 лет), а YFull записывает их «возраст» как одинаковый, либо 720, либо 2880 лет, выбор датировки фактически произвольный. В итоге получаются несуразные датировки.
Примеры – в переходе R1a-Y35 > YP7278 зафиксировано 9 промежуточных снипов (это приведено в таблице на сайте YFull), что соответствует 1300 лет разницы, однако, датировка для обоих снипов, родительского и нисходящего, приведена как одинаковая, 4300 лет назад. В переходе CTS3402 > CTS2613 зафиксировано 17 промежуточных снипов (2450 лет разницы), однако, датировка для обоих снипов в списке YFull – одинаковые 4300 лет назад, вместо 1850 лет назад для нисходящего, как должно быть (и как дает более точная датировка по гаплотипам). И так в таблицах YFull повторяется десятки и сотни раз.
Приходится на этом столь подробно останавливаться, потому что в последнее время усиленно распространяется легенда, которую безграмотно пропагандирует Балановский, что расчеты по снипам – значительно более точные, чем любыми другими методами. Историки могут принять эту легенду за чистую монету, и опять повторится ситуация последних 15 лет, когда практически все расчеты популяционными генетиками, опубликованные в академических журналах, были кардинально искажены, с завышениями до 300-400%. Для расчетов на времена удаленные, как правило, более 5-10 тысяч лет назад, и тем более на 100-200 тысяч лет назад, когда число снип-мутаций исчисляется многими сотнями, расчеты по снипам действительно могут оказаться полезными, но вводимые постулаты, как равенство датировок для «параллельных» снипов, опять сводит эту пользу к нулю, если не к отрицательным величинам.
Так что я весьма критически отношусь к расчетам датировок по снипам, если они не подтверждены перекрестными расчетами с использованием других методов, например, по мутациям в гаплотипах. Но часто бывает, что другие методы непригодны, или нет соответствующих гаплотипов. Тогда приходится принимать то, что есть. Но часто бывает, что совпадение расчетов по снипам и гаплотипам вполне удовлетворительное. Это придает уверенность, что в принципе подход, основанный на числе снипов, правильный, но он нуждается в доработке.
Но, как мы видим, попгенетики опять в своем амплуа – Балановский уже провозгласил, что метод расчетов по снипам самый точный, и ненавистные ему расчетные подходы ДНК-генеалогии можно отставить. Это опять было провозглашено без какого-либо исследования, и единственная статья, в которой Балановский этот метод использовал, дала неверные результаты. Дело в том, что Балановский в своей манере выхватил из многих вариантов лишь одну скорость мутации – 122 года на снип, без малейшего ее обоснования или проверки, и опять получил неверные датировки (Underhill et al, 2015, 32 автора, Eur. J. Human Genetics).
Теперь обратимся к статье Балановского с 22 соавторами (PLOS ONE, 2015), которую он представляет в своей «дискуссии» наряду с образцовыми, в которой, как объявляет, определил константу скорости мутации, равную 0.78х10-9 мутаций на нуклеотид в год. Надо же, и точность примечательная, с надежностью до второго знака, то есть почти 1%, что, конечно, не бывает и быть не может, но он так объявляет. И на диаграмме в своей «дискуссии» он своё значение помещает прямо в яблочко, остальные светила зарубежной попгенетики гуляют со своими значениями вокруг, а «наша – лучше всех».
Зная уровень Балановского как ниже среднего, я позволил себе усомниться, и данные пересчитал. И что получилось? Балановский взял данные по девяти казахам-аргынам, общий предок которых (по легендам) упомянут в хрониках в 1405 году, набросил 50 лет, выбрал из упомянутых девяти казахов шесть, привел их средний год рождения, и записал, что от общего предка до их современных шестерых потомков прошло 606 лет – замечаете, как работают попгенетики? Они возятся с мелочами, развивая якобы бурную деятельность, чтобы на «настоящую науку» было похоже, но пролетают по-крупному там, в чем безграмотны. В итоге получается пшик. Так и здесь, после всей этой возни с единицами лет Балановский написал, что среднее число снип-мутаций у шестерых выбранных казахов от их общего предка равно 4.67.
Когда я вижу, что среднее число снип-мутаций указывают с точностью до третьего знака, у меня, как у известного персонажа Пелевина, рука тянется к пистолету. Те, кто так «подают данные», либо жулики, либо безграмотные. Либо оба в одном флаконе. Смотрим на данные. Вот – числа мутаций от «общего предка» до каждого из шести современных потомков: 5, 5, 4, 7, 5, 2. Любой, имеющий элементарное арифметическое образование должен знать, что среднего из этого набора с точностью до третьего знака (то есть фактически до 0.1%) быть просто не может. Среднее здесь равно 4.67±1.11 без округления, с натяжкой – 4.7±1.1, а на самом деле 5±1. Но пусть пока без округления, тогда средняя скорость мутаций равна не 0.78х10-9, как объявляет Балановский, а (0.77±0.18)х10-9 мутаций на нуклеотид в год, или между 0.59х10-9 и 0.95х10-9, то есть в том же интервале, где эти определения были еще несколько лет назад, то есть между 0.6 и 1.0 миллиардных долей. И нужно ему было считать годы рождения казахов-аргынов с точностью до одного года? Мог спокойно им всем по сто лет поставить, и ничего бы в его расчетах не изменилось. Ничего нового Балановский со своим коллективом в уточнение константы скорости мутации не внес. Просто еще раз продемонстрировал вопиющую математическую безграмотность. И, конечно, полное отсутствие научной школы, что, впрочем, давно было ясно.
Переходим к следующему разделу. После всех его провалов по всему тексту, забавно, что он еще чего-то там «обсуждает», и называет это «научные дискуссии». Но раздел небольшой, всего шесть маленьких абзацев. Рассмотрим все шесть. Первый – пустой, кроме того, что трехкратные отличия «скоростей Животовского» от более приличных и обоснованных констант скоростей мутаций стали, по его мнению, «предметом оживленных дискуссий». Типичный стиль Балановского, что-то сказать, но вопрос не решить, и даже не попытаться.
Второй абзац – совершенно невразумительный и неверный. Балановский пытается совместить несовместимое, и цитирует Животовского, что мутации в гаплотипах возникают, конечно, с «генеалогической» скоростью, но часть гаплотипов «элиминируются дрейфом генов» (причем здесь гены-то? Опять Балановского в «гены» понесло, говоря о гаплотипах Y-хромосомы, в которых никаких генов нет). И, мол, потому константа скорости уменьшается, становясь 0.00069 мутаций на маркер за 25 лет. Это, конечно, глупость. Во-первых, как это «элиминирование» вдруг остановилось именно на константе 0.00069, и после этого, выходит, «элиминирования» больше не было. Во-вторых, «элиминирование гаплотипов» уменьшает количество как гаплотипов, так и мутаций в них, но среднее число мутаций на гаплотип продолжает сохраняться, на то это и константа скорости мутации. Серьезным случаем «элиминирования» является «бутылочное горлышко» популяции, но после его прохождения, если прохождение состоялось, мутации в потомках появляются опять согласно константе скорости мутации в маркерах и гаплотипах. Балановский всё не может понять, что «скорость Животовского», она же «популяционная скорость», является результатом некорректного анализа выборок гаплотипов. Они, эти выборки, являются смесью разных ветвей, и их нельзя подвергать расчету «чохом», как нельзя рассчитывать «время полураспада» смеси разных радиоактивных изотопов. А это фактически то, что делают в попгенетике.
Во всех тех примерах, которые Животовский выдавал за «калибровку», никакого «элиминирования» не было. Было манипулирование с выбрасыванием гаплотипов и подгонка под нужный ответ. А на самом деле и полинезийцы, и цыгане (см. выше) имели хороший набор гаплотипов, из которых вообще ничего не нужно было выбрасывать. Было 37 гаплотипов полинезийцев в 10-маркерном формате, во всех гаплотипах 49 мутаций от базового гаплотипа, среди всех 37 гаплотипов было 8 базовых гаплотипов. Четыре гаплотипа выпадали из общей серии, как показывает дерево гаплотипов (см. Клёсов А.А., «Происхождение славян», стр. 446). Даже если не выбрасывать те четыре гаплотипа, линейный метод дает 49/37/0.018 = 74 → 80 условных поколений, то есть 2000±350 лет до общего предка. Логарифмический метод дает [ln(37/8)]/0.018 = 85 → 93 условных поколений, то есть 2325±850 лет. Если же снять те четыре выпадающих гаплотипа, то логарифмический метод даст [ln(33/8)]/0.018 = 79 → 86 условных поколений, то есть 2150±790 лет. Мы видим, что все эти датировки совпадают в пределах погрешностей расчетов.
Подобная же ситуация с гаплотипами цыган, которых было 179 человек. Если считать по 8-маркерным гаплотипам, то линейный метод дает 59/179/0.013 = 25±4 условных поколений до общего предка, логарифмический метод дает [ln(179/26)]/0.013 = 27±4 поколений. Если считать по 6-маркерным гаплотипам, то линейный метод дает 34/179/0.0074 = 26±5 условных поколений до общего предка, логарифмический метод дает [ln(179/146)]/0.0074 = 27±4 поколений. Как видим, все четыре датировки практически одни и те же. Какое «элиминирование», с которым Балановский неуклюже пытается выручить «популяционную скорость»… Ясно, что Животовский считал совершенно неправильно, произвольно выбрасывал гаплотипы и подтягивал датировки.
А Балановский пишет – «Иного объяснения трехкратным различиям скоростей в научной литературе предложено не было, и объяснение элиминацией части гаплотипов вошло в обиход». Здесь и незнание, и ложь в одной фразе. Во-первых, там не «трехкратное» различие, а разнообразное. У 6-маркерных гаплотипов средняя скорость мутации 0.00123 на маркер за 25 лет, то есть отличие от «популяционной скорости» в 1.8 раз, у 17-маркерных гаплотипов средняя скорость мутации 0.0365, то есть отличие от «популяционной скорости» в 3.1 раз, у 25-маркерных гаплотипов – в 2.7 раз, у 37-маркерных гаплотипов – в 3.5 раз. Что касается того, что объяснения предложено не было – это ложь, я об этом подробно писал еще в 2009 году в журнале Human Genetics, там вся статья на шести страницах посвящена «методу Животовского».
Третий абзац в этом разделе Балановского – «оставалось непонятным, какой все же скоростью пользоваться на практике». Это только Балановскому и его соратникам подобного же низкого уровня квалификации оставалось непонятным. Подробные описания того, какой скоростью пользоваться, были даны в 2008 году на русском и английском языках в Вестнике, и активно обсуждались на сайтах, в частности, RootsWeb. В 2009 году все методы и расчеты были опубликованы в журналах Human Genetics, J. Genet. Geneal., Nature Preceedings. В 2010 году это было описано в книге «Происхождение человека». В 2011 году вышла статья совместно с И.Л. Рожанским в Advances in Anthropology, и с тех пор вышло еще много статей, последняя в 2014 году совместно с итальянским коллективом (Tofanelli, Francalacchi и др.). Впрочем, Балановский, напоминаю, писатель, а не читатель.
Четвертый абзац – опять ложь у Балановского. Он пишет – … данные полного секвенирования Y-хромосомы: в них «эволюционная» скорость не отличается существенно от «генеалогической». Примеров он опять не дает, как всегда, да таких примеров нет и быть не может.
Пятый абзац – опять Балановский запутался со своим стремлением все-таки протолкнуть «эволюционную скорость». Если попгенетикам – не жалко, им уже ничего не поможет. Он пишет, что «для более древних гаплогрупп «эволюционная скорость подходит лучше… примерно с возрастов порядка 30 тысяч лет». Это примерно как защищать остановившиеся часы тем, что все-таки дважды за сутки они показывают правильное время. Не подходит «эволюционная скорость» ни в каких случаях, просто считать надо правильно. На временах «порядка 30 тысяч лет» гаплотипы с «быстрыми» маркерами неприменимы, они начинают насыщаться возвратными мутациями, и число мутаций прогрессивно недосчитываются. Потому и кажется, что «скорость мутации» ниже, и при определенных временах она понизится в 3-4 раза, и приблизится к якобы «популяционной скорости». Но при еще больших временах она понизится еще больше, станет более низкой, чем «популяционная», и так далее. Нельзя же так, Балановский, свою безграмотность так выставлять напоказ. Стыдно должно быть. И это он вставляет в свою книгу! Да, хороший подарок готовится попгенетикам… Кто же такой мусор публиковать будет? Какое издательство? В глаза бы этим издателям посмотреть.
Последний, шестой абзац. Там – гроздь банальностей и непонимания вопроса. Начинается с того, что «окончательной ясности в этом вопросе нет» (то есть в расчетах времен до общих предков по гаплотипам). Ясность там давно есть, просто учиться надо. Читать толковые статьи и книги, где все объяснено. Здесь же у Балановского «возраст гаплогрупп теперь достаточно надежно определяется по результатам полного секвенирования Y-хромосомы». К сожалению, это пока далеко не так, и это описано выше.
Чем дальше по «новой статье» Балановского, тем более это похоже на хрестоматийное описание Ильфом и Петровым рукописи композитора Листа, где на первой странице указано играть быстро, а далее «гораздо быстрее», «быстро как только возможно», и потом «еще быстрее!». Только у Балановского «глупо», «гораздо глупее», «глупо как только возможно», и далее «еще глупее». В этом – весь Балановский. Его следующий раздел озаглавлен «Околонаучные и лженаучные дискуссии». Его даже разбирать неинтересно. Представляете – Балановский судит (!) и решает, какие дискуссии «околонаучные», и какие «лженаучные». Интересно, зеркало у него есть?
Надо сказать, что систематическая порка Балановского в этом моем разборе его позорного «произведения» – для меня вовсе не самоцель. Я Балановского порол уже столько раз, что уже давно неинтересно. Цель этого разбора – образовательная, преподавательская. Это, если угодно, мини-учебник. Я даю определения, описания методов, примеры расчетов, то, чего у Балановского нет и близко. Его наивные и примитивные описания, что он считает «околонаучным» и что «лженаучным» – это просто смех. Там и разбирать нечего, особенно после моих объяснений и описаний выше. Там, в этом разделе Балановского, специалисту делать нечего. Балановский все время пытается вывернуться, выскользнуть, поскольку он уже, видимо, начал понимать, что что-то у него не так, причем сильно «не так». Он уже пишет, что считать датировки – это не важно, это вопрос незначительный.
Ну что мне, положить перед ним толстую кипу его статей, в которых датировки были принципиально неверными, и неверными были «интерпретации исторического характера»? Положить статьи его коллег-попгенетиков, которые датировали общих предков ариев в Индии 12-16 тысяч лет назад? Статьи Балановского, которые датировали субклад R1a-M458 (в основном славянский) временами 10 тысяч лет назад, и пришли к выводу, что арии в Индию не приходили? И на основании этой статьи Л. Клейн написал свой мусор, показав полное отсутствие исторического чутья. Статьи коллег-попгенетиков Балановского, которые датировали расхождение христиан и мусульман на Ближнем Востоке временами 4 тысячи лет назад, когда правильные датировки дают 7-8 век н.э., время начального распространения ислама? Это же форменный цирк… И такого мусора в академической литературе – масса. А Балановский, изворачиваясь, пишет – «трудность в определении абсолютного возраста гаплогрупп не скажется существенно на выводах». И здесь до него не доходит, что ерунду он пишет. А, скорее всего, доходит, но преобладает желание извернуться, не признаться, что фатально неправ.
Забавно, что Е. Балановская демагогически восклицала, что я богатство популяционной генетики называю «мусором». Да, действительно яблоко от яблони недалеко падает. Какое богатство? То, что Балановский несет в своей «дискуссии» – это богатство? Позор это, а не богатство. Вопиющая безграмотность.
Балановский «объясняет», что «представителям науки» неинтересны дискуссии на форумах, и удивляется, что там активно критикуется «эволюционная скорость». Да, она критикуется уже немало лет, причем критикуется предметно, с приведением конкретных примеров ее некорректности, что у Балановского, «представителя науки», как он себя именует, полностью отсутствует. Как-то на англоязычном форуме было сообщение венгерского популяционного генетика, которого подняли на смех, что в недавней статье с его участием была использована «популяционная скорость» и, естественно, получены абсурдные выводы. Он извинился и рассказал, что поначалу авторы использовали нормальные расчеты, с «генеалогической» скоростью, но рецензенты и журнал поставили условие, что если не будет использована «популяционная скорость», то статья будет отклонена. Авторы смалодушничали, и заменили расчеты на абсурдные. Участники дискуссии это осудили как измену науке, что ученый так поступать не должен. А Балановский был одним из тех, кто эту измену науке поддерживал и пропагандировал. И он называет себя «представителем науки»!
Вот где подмена понятий в исполнении Балановского. И сейчас в том же духе он изворачивается, и называет выбор констант скоростей мутации вопросом непринципиальным (!). При этом посвятил его обсуждению 14 страниц своей «дискуссии», этому «непринципиальному вопросу». Нет, он не представитель науки. Вот в чем драма Российской академии наук – лабораторией в ее стенах, в Институте генетики, руководит Балановский, человек безграмотный, изворачивающийся, лживый, недалекий.
В этой части «дискуссии» у Балановского опять недоразумение громоздится на безграмотность. Он пишет – «Любопытно…, что многие генеалоги начисто отрицают эволюционную «скорость Животовского», но при этом пользуются методом ASD», и при этом дает ссылку на (Sengupta et al, 2006), напирая на то, что метод разработан Животовским. И здесь Балановский показывает незнание предмета. Метод ASD (average squared difference, или как вариант average squared distance) не имеет никакого отношения к «скоростям Животовского». Это общепринятый в математической статистике метод расчета средних квадратичных отклонений, применяемый в различных вариантах. В ранних работах популяционной генетики его применяли (Goldstein, Linares, Cavalli-Sforza и Feldman, PNAS, 1995; Goldstein, Linares, Cavalli-Sforza и Feldman, Genetics, 1995), но это была не «разработка» метода, а его применение к описанию вариации в маркерах гаплотипов. Об этом и говорят названия статей того времени – «Оценка генетических расстояний из микросателлитных участков» (Goldsein et al, 1995, Genetics) или «Генетические расстояния и реконструкция филогенетических деревьев из микросателлитных ДНК» (Takezaki и Nei, 1996, Genetics). Причем здесь статья Sengupta через десять лет? Но главное то, что «эволюционная скорость» здесь совершенно не при чем, о чем, Балановский, видимо, не имеет понятия. В методе ASD, который для простоты назван в моих статьях и книгах «квадратичным», по формуле матстатистики рассчитывается квадрат разницы в числе аллелей попарно в каждой аллели и всеми другими аллелями, суммируется по определенным правилам и делится на число маркеров, на квадрат числа гаплотипов и на константу скорости мутации на маркер.
Например, имеем серию из трех 12-маркерных гаплотипов
13 24 16 10 11 15 12 12 10 13 11 30
13 25 16 11 11 15 12 12 10 13 11 30
13 24 16 11 11 15 12 12 11 13 11 30
Производится расчет разницы в аллелях «всех между всеми». В данном случае сумма квадратов их отклонений друг от друга равна 12 + 12 + 12 + 12 + 12 + 12 + 12 + 12 + 12 + 12 + 12 + 12 = 12, деленная на 2, поскольку в каждом маркере число квадратов мутаций считалось дважды, деленное на число маркеров в гаплотипе (12) и на квадрат числа гаплотипов (9), и, наконец, на величину константы скорости мутации на маркер для 12-маркерных гаплотипов (0.00167). Получаем 12/2/12/9/0.00167 = 33 условных поколений, то есть 825±480 лет до общего предка. Погрешность рассчитана по стандартным формулам, которые я много раз показывал в своих работах. Она в данном случае относительно велика, поскольку в этих трех гаплотипах имеются всего три мутации. В квадратичных методах расчетов поправка на возвратные мутации не вводится, она уже учтена самим методом расчета.
Ну, и как понять этот безграмотный пассаж Балановского – «многие генеалоги начисто отрицают эволюционную «скорость Животовского», но при этом пользуются методом ASD»? Впрочем, его и понимать не нужно. Не имеет Балановский понятия, но при этом пишет книгу. Да, «писатель». И будут по этой ерунде учиться новые поколения попгенетиков. Если, конечно, не будут приняты разумные меры со стороны РАН. Я готов в этих мерах оказать содействие.
Вершиной «творчества» Балановского является раздел «Лженаучные дискуссии», практически полностью посвященный мне. Это тот самый случай, когда для Балановского «назло маме уши отморожу». Опустим его безграмотные общие рассуждения о «лженауке», это мало кому интересно. Перейдем к сути раздела. В нем – семь пунктов. Опять, опустим словоблудия Балановского, перейдем к выводам самого Балановского по каждому пункту.
Пункт 1 – о скоростях мутирования в гаплотипах. Вывод – мои подходы «вряд ли приведут к научным ошибкам», поэтому они «не вызывают нареканий».
Пункт 2 – о логарифмическом методе. Вывод – «и здесь особых проблем нет… остается в рамках правомочных… методов».
Пункт 3 – что я якобы «объявил «генеалогическую скорость» своим изобретением». Опять глупость, и понятно, почему Балановский обходится здесь без цитат, когда и где я это «объявил». Понятно потому, что он сам это придумал. Не «генеалогическую скорость» я чем-то там объявил, я это понятие вообще не использую. Нет никакой «генеалогической скорости», есть константы скоростей мутации, свои для гаплотипа каждого используемого формата. Я уже приводил примеры – для 6-маркерного гаплотипа она равна 0.0074 мутаций на маркер на условное поколение в 25 лет, для 12-маркерного гаплотипа – 0.02 (в той же размерности), для 17-маркерного гаплотипа 0.0365, для 25-маркерного 0.046, для 37-маркерного 0.090, для 67-маркерного 0.12, для 111-маркерного 0.198 мутаций на гаплотип на условное поколение. Если кто захочет узнать средние значения констант скоростей в расчете на маркер, то следует разделить приведенные выше значения на число маркеров, получим соответственно (х10-3) 1.23, 1.66, 2.15, 1.84, 2.43, 1.79, 1.78. Ну и какая именно из них «генеалогическая скорость мутации»? Это настолько же глупо, как называть константы скоростей бесчисленного количества химических реакций все чохом «скоростями реакций», придавая этому некий количественный смысл. Опять безграничная безграмотность Балановского.
К тому же он пишет – Если А.А. Клёсовым и была независимо подтверждена известная задолго до него и многократно подтвержденная генеалогическая скорость… – она остается все той же… «генеалогической». Что это за «генеалогическая скорость», кем и как она была «многократно подтверждена» – Балановский, конечно, умалчивает. Подскажем ему – несколько раз в литературе была использована константа скорости мутации 2.0х10-3, которая отличается от всех, приведенных выше в этом абзаце, и она использовалась, как принято в попгенетике, одна на все случаи. Но это было только на раннем этапе попгенетики, начиная с 2004 года она была затоптана ложной «эволюционной скоростью», которую только и принимали рецензенты генетических журналов. Исключением была только моя статья в Human Genetics, которую, кстати, приняли к печати ровно через неделю после представления в журнал.
Только в 2011 году Балановский опомнился, и решил сравнить «эволюционный» и некий «генеалогический» метод, принимая к 17-маркерным гаплотипам константу скорости мутации 0.0021 на маркер на поколение в 30 лет, ссылаясь на работы по парам отец-сын. Строго говоря, так делать нельзя. Не было «30 лет на поколение» в парах отец-сын. Величина 0.0021 была получена в расчете на поколение, а не на 30 лет. Введение 30 лет без калибровки – это произвол, в науке не допустимый. При этом из расчетов Балановского удалялись маркеры DYS385a и DYS385b, так что фактически использовались не 17-, а 15-маркерные гаплотипы. Тем более если величина 0.0021 по парам отец-сын на самом деле ближе к поколению в 25 лет (мы этого просто не знаем), то при переводе в 30 лет на поколение константа станет уже 0.0025 мутаций на маркер. Коротко говоря, это все топорно. Вовсе не академично. Резюмируем – про мое «изобретение генеалогической скорости» – это опять отмораживание ушей у Балановского назло его маме. Не было этого у меня – зачем Балановскому так подставляться?
Пункт 4 – о «пошаговой инструкции расчета возраста», как я понимаю, о моей таблице по расчету возвратных мутаций (Вестник Академии ДНК-генеалогии, 2008, том 1, № 5, стр. 812-835). Балановский ссылок опять не дает. Но он пишет – «вещь, несомненно, полезная», «справочная таблица… тоже полезна», «итак, и на этом этапе противоречий… нет».
Пункт 5 – опять о поправках на возвратные мутации. Балановский пишет – «поправка… правильна» (ай, спасибо, хотя похвала от плохенького середнячка никакой ценности не имеет). Дальше он опять переходит на бессмыслицы, типа «ведь обратные мутации могли происходить и в примере, послужившем основой калибровки». Опять Балановский не имеет понятия, о чем пишет. Какой пример, который «послужил основой калибровки»? Как водится, Балановский просто вбрасывает какие-то слова. Для калибровки используются данные документальной генеалогии, какие там «обратные мутации»? Далее Балановский пишет – «итак, и тут в методе расчета ДНК-генеалогии нет ошибок? Возможно». Ну и замечательно, Балановский хоть начал что-то понимать.
Пункт 6 – это уже поток сознания у Балановского. Он начинает оспаривать положение, им самим придуманное, что мои методы расчетов являются «единственно верными». Цитат, конечно, опять нет. И быть не может. Потому что в науке нет ничего «единственно верного». И как он оспаривает то, что сам придумал? Что эти методы, во-первых, уже давно «разработаны популяционными генетиками», во-вторых, что они «не единственно верные» и «имеют свои недостатки», «да и «генеалогическая» скорость мутирования не бесспорна». Про «генеалогическую скорость» я уже говорил, я этот термин не использую, да и нет такого. Поэтому я не знаю, что там «не бесспорно», когда сам термин неверен. Или он про константы скоростей мутаций гаплотипов и маркеров, которые я приводил выше? Понятия «бесспорны» или «небесспорны» к ним вообще не применимы, это от безграмотности Балановского. Они оптимальны на этапе нашего современного знания. Знание продвинется, и константы могут измениться. Я их сам подправляю время от времени, когда накапливается больше данных.
Пункт 7 – это то самое «еще быстрее», о чем я иронически писал выше. Балановский подводит итог предыдущих шести пунктов, и заключает, что у меня «лженаука». Ну, не клоун ли Балановский? В четырех пунктах был вынужден признать, что мои подходы верные, в двух пунктах что-то сам придумал и приписал мне, что я якобы что-то не сам изобрел, а это было давно известно в попгенетике – так что там тогда «лженаука»? Попгенетика?
А ничего, это стиль Балановского. Те самые инсинуации, совершенно безосновательные. Просто у него другого выхода нет, как повторять «лженаука», авось кто-нибудь клюнет. Но если и клюнут, то маргиналы. Напротив, вот несколько выдержек из писем читателей за последние несколько дней, в дополнение к той, с чего началась эта статья:
>> Уверенность в правильности Ваших расчетов все больше возрастает. Очередную статью Балановского «Дискуссии о датировках» иначе как инсинуациями не назовешь.
>> Если обыватель еще и почитает хотя бы некоторые Ваши статьи, все становится очевидным, что Балановский просто лжец.
>> После прочтения серии других статей Балановского, которых у него немало в последнее время, складывается впечатление, что Балановский либо некомпетентен и вообще не читал Ваши книги и статьи, либо реально тупо руководствуется принципом «лишь бы возразить».
>> Хорошо, что Балановский признал правильность генеалогических скоростей. Некуда деваться… Опять же маленько соврав по поводу того, кто эти скорости калибровал и кому первому принадлежит авторство их публикации. Я знаю, что Вы, все знают, это все давно опубликовано и в соответствующих ссылках Ваших статей это все есть. Но, мне кажется, после этой статьи Балановского нужно еще раз напомнить.
Вот об этом – настоящая статья. Балановский такой статьи не стоит, он здесь выполняет роль боксерской груши. Статья, как я уже упомянул выше – мини-учебник по константам скоростей мутаций и сопряженным вопросам. Хотелось бы, чтобы ее так и воспринимали.
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке
| Переформат.ру пока находит силы держаться, но вопрос выживания сайта по-прежнему стоит очень остро. Чтобы продолжить «общественно-научное переформатирование» мы решили запустить проект на «Планете». Первые успехи были заслугой сравнительно небольшого круга наших читателей и сторонников, за что им сердечное спасибо (и презенты). Поэтому вопрос снова подвис. Даже от минимальной суммы, необходимой для продолжения Переформата, мы пока далеки. Переформат.ру: будет ли продолжение?.. |

















