
Лидия Павловна Грот недавно побывала в Воронеже на презентации своей вышедшей в прошлом году книги Прерванная история русов: Соединяем разделённые эпохи. В выступлении она рассказала о том, каким важным инструментом в ее исследованиях оказалась ДНК-генеалогия, и сообщила, как сейчас представляется картина миграций далеких предков гаплогруппы R1a, которую сейчас имеют в среднем половина этнических русских. Ниже – выдержка из письменного варианта тезисов ее сообщения.

Наш род или гаплогруппа R1a. ДНК-генеалогия называет его праславянами или славянами, т.к. к нему принадлежит большинство современных славян. Его представители примерно 4900-4600 лет тому назад передвинулись на Среднерусскую равнину, шли и с Балкан, и с Южной Балтии, с территории современной Германии как бы полосой. Примерно 4500 лет назад они стали расходиться по разным направлениям как легендарные арии – на юг, через Кавказ в Месопотамию, на Ближний Восток (митаннийские арии) и Аравийский полуостров; на юго-восток, в Среднюю Азию и далее, через 500 лет, то есть примерно 3500 лет назад – на Иранское плато (авестийские арии).
После ухода ариев на восток примерно 4500 лет назад в Восточной Европе осталась ветвь R1a-Z280, к которой относится большинство современных этнических русских, соответственно, по моему предложению, с которым согласился А.А. Клёсов, эта ветвь представляла древних русов. Вот эти древние русы, а также та часть ариев, которая осталась на Русской равнине и влилась в состав русов, стали наиболее древними предками русских, украинцев, белорусов. Иначе говоря, русские имеют с ариями одних и тех же предков, которые с распадом индоевропейской общности в течение III тыс. до н.э. разошлись, как расходятся сыновья одного отца. Поэтому, как подчеркивает А.А.Клёсов, «славяне», «арии», «скифы» это в своей основе – одни и те же люди, один род, но разных исторических эпох. Они связаны прямой наследственностью в рамках рода R1a. Вот начальный рубеж нашей истории, от него мы должны идти…
Я бы внес некоторые коррективы, а именно что носителей R1a можно называть славянами только в определенном контексте, только в применении к современным носителям языков славянской группы – чехам, полякам, болгарам, сербам, словенцам (и другим жителям бывшей Югославии), белорусам, украинцам, русским и другим, попадающим в эту категорию. Скажем, ирландцев или бельгийцев, уйгуров или индийцев, у которых найдена гаплогруппа R1a, славянами назвать никак нельзя. Если первых относительно мало в сравнении с носителями гаплогруппы R1a в восточной Европе, то индийцев с гаплогруппой R1a уже примерно 100-200 миллионов (мужчин, естественно), по разным оценкам. Они – не славяне. Если же говорить об их предках, то их можно условно назвать праславянами, если договориться об определении, кто такие были праславяне.
Скажем, далекие предки тех же ирландцев или бельгийцев с гаплогруппой R1a жили в основном на Русской равнине, многие имели субклад R1a-Z280, как и большинство этнических русских гаплогруппы R1a, и их вполне можно было, на мой взгляд, назвать праславянами. Они пришли, в конце концов, на Британские острова и в Центральную Европу с Русской равнины – кто мирной миграцией, кто боевым путем (как скифы, например, или римские легионеры), кто попал в плен или осел там другими путями. Далее, славяне – это не только R1a, которых примерно половина русских, поляков, украинцев, белорусов, треть сербов, хорватов, словенцев, но это и носители гаплогрупп I1, I2, N1c, и многих относительно немногочисленных гаплогрупп, называющих себя этническими русскими или другими народами славянской группы. Называть славянами только носителей R1a – это обижать других.
В общем, это относительные мелочи, и они происходят от неустоявшихся определений. В основном же, тезисы Л.П. Грот правильны, и продвигают вперед историческую науку. Будут появляться новые данные, вноситься уточнения, как обычно в науке, но на сегодня это вполне разумная концепция.
Это была затравка, не об этом я хотел поговорить. А о том, с какими трудностями, в основном, ментального характера (а вовсе не научного) оппонентов продвигаются положения ДНК-генеалогии в массы, далекие от критического осмысления того, что они слышат. Именно с такими оппонентами встретилась Л.П. Грот в Воронеже. Заметьте, что основной тезис оппонентов, о которых я сейчас говорю – не возражения научного характера, не противопоставление фактов, которые бы противоречили данным ДНК-генеалогии, не прочая научная аргументация. Это – тупое «а Баба-Яга против». Другими словами, «а вот другие полагают иначе», где другие – это неназванные «популяционные генетики».
Здесь надо сказать, что я еще ни разу не встречал популяционных генетиков, которые бы мне возразили аргументированно, с выставлением контраргументов, показали, в чем мои расчеты неверны, показали, как нужно считать и почему их расчеты лучше. На это есть простая причина – они сами вовсе не уверены, что считают правильно. Они не разбираются. Они знают, что так принято; как им предписано считать, они так и считают. Почему, на чем это «так принято» основано, они не знают, а если и слышали, то никогда критически в этом не разбирались. Вот это и есть результат отсутствия у попгенетиков научной школы. То, без чего наука погибает. Она, наука, у них в огромной степени уже погибла, и это последовательно происходило последние десять лет, со времени узаконивания у них в 2004 году «популяционных скоростей мутаций», или «скоростей Животовского». Я об этом расскажу ниже.
Надо сказать, что Л.П. Грот была в восторге от тех, кто ее пригласил и организовал дискуссию. По ее словам, это отличные люди, энтузиасты русской истории и культуры, современные и прагматичные. Приятно было, по ее словам, то, что дискуссия собрала много участников, особенно приятно было, что пришло много преподавателей воронежских вузов – историков, юристов, археологов. Правда, оказалось, что для археологов исследования Л.П. Грот о норманизме как части шведского политического мифа были мало известны, поэтому ими высказывались взгляды, не выходившие за рамки классического норманизма, однако, был и явный интерес к выступлению, а это всегда – вероятность продолжить разговор, что как раз и требуется от подобных встреч. Но на дискуссию пришли также те, для которых «а Баба-Яга против». Среди них и воронежский специалист по историографии «Ригведы», некто А. Семененко. В генетике он, естественно, не разбирается, и тем более в ДНК-генеалогии… в общем, мы ниже предоставим ему слово. На презентации Л.П. Грот он высказывался теми же словами, что и в своем блоге. Блог я не собираюсь рекламировать, кому надо, найдут, но цитаты из него дам. Мне этот Семененко, разумеется, безразличен, как и его мнение, но его слова показательны. Их полезно разобрать, именно аргументированно. Одно дело мне излагать положения в статьях, но многие в статьи, как показывает опыт, «не врубаются», или просто их не читают, и тем не менее выскакивают с критическими замечаниями. Поэтому статьи часто не помогают многим разобраться в вопросе, и вот тогда полезны разборы слов «критиков».
«Критикам», как правило, это не помогает. Они же не «истину» ищут, для них важен злобно-критический процесс (ниже мы увидим слова типа «бред»), они этим живут. Рецепторы здравомыслия у них отсутствуют, те места другими рецепторами заняты. Но вот другим, кто своего мнения пока не имеет, эти контраргументы порой полезны.
Итак, в блоге Семененко один из участников пересказывает (не очень точно, но приемлемо) мои слова, говоря о гаплогруппе R1a – «у половины русских есть эта гаплогруппа, но она у них появилась 5 тыс. лет назад, а у арабов 3,5 тыс. лет», и далее – «профессор Клёсов говорит на 25-ой минуте, что арии появились на Балканах и стали двигаться на восток. Вроде бы это расходится с вашими данными, или нет».
Далее отвечает Семененко. Заметим, что никакого ответа по сути заданного вопроса он не дает. Люди без тех самых рецепторов на прямые вопросы не отвечают, им это ни к чему. Более того, для ответа надо знать материал, разбираться в нем. А они не знают и не разбираются. Поэтому смотрите, так сказать, за руками того, кто отвечает. Поскольку Семененко вываливает ворох несуразиц, и на вопросы не отвечает, его пассажи я буду комментировать по частям, для ясности.
Итак, приступаем…
Семененко: Профессор Клёсов — не генетик. Я опираюсь на данные международных коллективов профессиональных генетиков, опубликованные в ведущих международных научных журналах по генетике. А профессор Клёсов у них пользуется очень дурной славой. Он считает, что разработал собственную методику определения скорости накопления генетических мутаций, поэтому его «датировки» расходятся с датировками профессиональных генетиков.
Заметим опять, что, как принято у людей без того самого рецептора, разговор переключается на другое, ответа на заданный вопрос не дается. Вместо этого почему-то приплетается, что я не генетик. Я и не нейрофизиолог, и не специалист по глотанию шпаг. Но при чем здесь генетика? Я – специалист по скоростям химических и биологических реакций, автор книг и университетских учебников по этой тематике, кстати, удостоен Госпремии СССР по науке именно за работы по скоростям и константам скоростей реакций, и здесь не имеет значения, это скорости накопления мутаций в ДНК или скорости других процессов, закономерности, в целом, одни и те же. Именно знание этих закономерностей и заставило меня ужаснуться тому, как это делают популяционные генетики. А они следуют «методу», который разработал Л.А. Животовский с коллегами десять лет назад, применив совершенно искаженные представления и безумные допущения.
Для начала отметим, что Л. Животовский применил совершенно усредненные и искусственные приближения, приняв, что константа скорости мутаций в Y-хромосомах ДНК должна быть усреднена по 10 тысячам (!) гаплогрупп за все время существования человечества, и тогда у него получилось, что она, константа скорости мутации, должна быть равна 0.00069 мутациям за 25 лет на маркер, причем она одинакова для любых гаплотипов, 6-, 7-, 8-, 9-, 10-, 12-маркерных и так далее. Любой, кто имеет хотя бы минимальный опыт работы с гаплотипами знает, что для каждого маркера есть своя константа скорости мутации, и в 6-маркерные гаплотипы попадают одни маркеры, в 8- или 9-маркерные свои (и, значит, средняя константа скорости мутации будет уже другой), в 10- или 12-маркерные гаплотипы добавляются еще другие маркеры со своими константами, и так далее. Считать, что константа одна на все – это вносить значительную погрешность в расчеты. Именно эту ошибку делают все без исключения «международные коллективы профессиональных генетиков», чьи фундаментально ошибочные статьи «опубликованы в ведущих международных научных журналах по генетике». Это – трагедия современной популяционной генетики. А люди без тех самых рецепторов, не думая, не зная и не понимая, продолжают кудахтать, другого слова не подберу, что «он не генетик». Это-то здесь при чем? Нет в ДНК-генеалогии генетики, генетика закончилась до ДНК-генеалогии, когда генетики определили, какие мутации есть в ДНК, сколько «тандемных повторов» есть в маркерах, и какие снипы найдены в каких участках Y-хромосомы. Иначе говоря, генетика, а точнее, лаборанты в компаниях типа FTDNA, снабжают нас этими данными, а ДНК-генеалогия идет дальше, проводя количественную обработку этих данных, чего генетики делать уже не умеют. У них образование не то, их этому не учили. А учиться, как показывают последние 6-8 лет, они или не могут, или не хотят.
Давайте посмотрим, насколько промахиваются популяционные генетики, применяя свои «популяционные скорости». Как уже было сказано выше, для 6-, 7-, 8-, 9-, 10-, 11-, 12-, 17-маркерных гаплотипов (а с более протяженными популяционные генетики уже не работают, а если что-то и появилось в последнее время, то это как исключение) константа скорости мутации для всех равна 0.00069 на маркер на 25 лет, то есть для восьми типов гаплотипов выше суммарная константа скорости мутации равна, соответственно, 0.00414, 0.00483, 0.00552, 0.00621, 0.00690, 0.00759, 0.00828, 0.01173 мутаций на гаплотип. На самом деле, эти скорости равны, как показано на множестве примеров, в том числе откалиброванных по данным документальной генеалогии, 0.0088 (6-маркерные гаплотипы); 0.013 (7-маркерные); 0.013 или 0.0168 (в зависимости от структуры 8-маркерного гаплотипа – у одних пропущены маркеры 385а, 385b, 426 и 439, у других 426, 388, 439 и 392), 0.017 или 0.018 (в зависимости от структуры 9-маркерного гаплотипа, используемого разными авторами), 0.018 и 0.022 (у 10-маркерных), 0.019 и 0.020 (у 11-маркерных), 0.020 (в формате FTDNA) и 0.021 и 0.024 (в других форматах), 0.034 (в формате Y-filer).
Как видим, ошибка популяционных генетиков (в сторону завышения датировок) составляет, соответственно, 213% (для 6-маркерных гаплотипов), 269% (для 7-маркерных), 236-304% (для 8-маркерных), 274-290% (для 9-маркерных), 261-319% (для 10-маркерных), 250-264% (для 11-маркерных), 242-290% (для 12-маркерных), 290% (для 17-маркерных). Если продолжить этот ряд, то для 37-маркерных гаплотипов «популяционная константа Животовского» даст величину 0.00069х37 = 0.02553 мутаций за 25 лет, а настоящая, откалиброванная величина равна 0.090, то есть ошибка попгенетиков составит уже 353% в сторону завышения датировок. Если, например, арии пришли в Индию 3500 лет назад, то «константа Животовского» даст 12 355 лет. Так попгенетики и получают (см. например, статью Underhill et al, 2010), и это привело их к следующему выводу (цитирую заключительную фразу абстракта к цитируемой статье Underhill: Importantly, the virtual absence of M458 chromosomes outside Europe speaks against substantial patrilineal gene flow from East Europe to Asia, including to India, at least since the mid-Holocene.
То есть, на основании своих безумных расчетов они получили, что гаплогруппа R1a не могла придти в Индию ранее 12 тысяч лет назад (на самом деле они получили 14 тысяч лет назад), поэтому, по их заключениям, никакие арии в Индию не приходили, во всяком случае, до середины голоцена. Датировку R1a-M458 они «засандалили» во времена 11 200 лет назад (на самом деле, в три раза меньше), и заключили, что поскольку их в Индии нет, то опять же до голоцена никакие R1a в Индию не приходили. В действительности же, датировка R1a-M458 в Европе не превышает 4000 лет назад, а к тому времени арии уже ушли на юг и восток, потому-то R1a-M458 с ними не было, просто не успели.
И вот таким мусором заполнены академические издания в области попгенетики. В цитируемой статье такая обойма авторов: Underhill, Myres, Руутси, Metspalu, Животовский, King, Lin, Chow, Semino, Battaglia, Кутуев, Ярве, Chaubey, Ayub, Mohyuddin, Mehdi, Sengupta, Рогаев, Хуснутдинова, Пшеничнов, Балановский, Балановская, Jeran, Augustin, Baldovic, Herrera, Thangaraj V. Singh, L. Singh, Majumder, Rudan, Primorac, Виллемс, Кивисилд. Представляете, 34 автора (российских и эстонских я выделил кириллицей, страна должна знать своих героев), и никто не понимает, что они делают, как и зачем так считают. В итоге полностью искаженное представление истории. Это же вынесено в виде критики на «мою» страничку в Википедии (хотя не я ее писал и представлял, там немало погрешностей в моей биографии), в которой делается вывод, что я неправ, и арии в Индию не приходили, и что это якобы современные представления. И дается галерея таких вот «мусорных» ссылок попгенетиков. Стыд и позор тем, кто такую ерунду написал, а редакция Википедии приняла.
Вот такая цена словам Семененко и иже с ним, что «Я опираюсь на данные международных коллективов профессиональных генетиков, опубликованные в ведущих международных научных журналах по генетике». Грош цена этим «международным коллективам профессиональных генетиков», а также рецензентам этих «международных научных журналов».
А профессор Клёсов у них пользуется очень дурной славой. Он считает, что разработал собственную методику определения скорости накопления генетических мутаций, поэтому его «датировки» расходятся с датировками профессиональных генетиков.
Теперь понятно, что за этим кроется, и почему «дурная слава». Не могут они ничего противопоставить, кроме как тихо ненавидеть. Когда я в далеком 2009 году разнес на 14 страницах в журнале Human Genetics их «подходы», они (тот же Животовский с коллегами) ответили в конце своего «ответного комментария – «Finally, regarding the detailed “haplotype trees” offered by the Comment, these are indeed interesting and can be very instructive». То есть, «это интересно и может быть очень инструктивным». За прошедшие пять лет, впрочем, этой «очень инструктивности» авторы не проявили. Вот их фамилии: Michael Hammer, Doron Behar, Tatiana Karafet, Fernando Mendez, Brian Hallmark, Tamar Erez, Lev Zhivotovsky, Saharon Rosset, Karl Skorecki. Так что, как видим, «международный коллектив профессиональных генетиков», не имеющих понятия, что они фундаментально ошибаются в своих подходах, отнюдь не маленький. Это – цвет международной популяционной генетики.
Дам еще иллюстративный пример. В Шотландии есть знаменитая «генеалогическая» семья МакДоналдов, их несколько тысяч человек, на них работает целый штат профессиональных генеалогов, их документальная генеалогия изучена во всех возможных деталях. И неудивительно, они берут свое начало от шотландского вождя Сомерледа, среди их предков наполеоновский маршал МакДоналд и много других выдающихся людей. Основная группа МакДоналдов гаплогруппы R1a ведет свою линию от Джона Лорда Островов (John Lord of the Isles), умершего в 1386 году, то есть 628 лет назад. Принимая 25 лет на поколение (именно эта величина входит в величины констант скоростей мутаций, приведенные выше), получаем, что Джон умер 25 поколений назад, то есть жил примерно 26 поколений назад. В своей книге «Происхождение человека» (2010) я рассмотрел серию гаплотипов потомков Джона, которая тогда насчитывали сначала (в 2008 году) 68 гаплотипов, затем (в 2010 году) 84 гаплотипов. Среди 68 гаплотипов в 6-маркерном виде было 17 мутаций, и 53 гаплотипа были немутированными, то есть базовыми. Считаем по правилам ДНК-генеалогии. Сначала по мутациям, используя так называемый линейный метод: 17/68/0.0088 = 28±7 поколений до Джона (погрешность велика, поскольку мутаций мало). Теперь считаем без учета мутаций, используя логарифмический метод: [ln(68/53)]/0.0088 = 28±5 поколений назад. 0.0088 – это константа скорости мутаций для 6-маркерных гаплотипов, см. выше. Как видим, наблюдается совершенно разумное совпадение с документальной генеалогией по Джону и его потомкам.
Считаем «по Животовскому»: 17/68/6/0.00069 = 60 поколений, то есть 1500 лет до Джона. Это называется «ни в какие ворота». Не говорите об этом расчете генеалогам семейства МакДоналдов, засмеют и выгонят, откажутся разговаривать.
В 2010 году из 84 гаплотипов 60 были 25-маркерными, в них 69 мутаций, из 60 гаплотипов 18 было идентичных, то есть базовых. Посчитаем по ним. Линейный метод: 69/60/0.046 = 25±4 поколений, логарифмический [ln(60/18)]/0.046 = 26±4 поколений назад (0.046 мутаций на гаплотип на условное поколение в 25 лет – это константа скорости мутаций для 25-маркерных гаплотипов). Как видим, и здесь хорошее совпадение с документальной генеалогией.
Считаем «по Животовскому»: 69/60/25/0.00069 = 67 поколений, то есть 1675 лет до Джона. Опять в те же ворота, то есть «ни в какие». Вот так работают расчеты «популяционных генетиков». Как видим, разница та же, завышение в датировке на 260-270%, в данном конкретном случае. А поскольку случаи разные, то попгенетики задирают датировки обычно в 2.5-4.0 раза. Вот такая цена их расчетам.
«Знающий» попгенетик возразит – не может быть, Животовский свои скорости мутаций калибровал, это все популяционные генетики знают, и основополагающая статья его 2004 года основана на калибровках его скорости мутации, той самой, 0.00069 мутаций на 25 лет на маркер. Там всё доказано, потому его константу и взяли за основу в популяционной генетике.
И вот здесь начинается самое интересное. Похоже, ни один популяционный генетик, ни один (!), не удосужился проверить сам, лично, что там была за «калибровка». Ни один не взял исходные данные Животовского и не проверил. Я это сделал. Давайте посмотрим, как Л. Животовский калибровал. Основная его «калибровка» – по группе болгарских цыганских таборов, 8-маркерные гаплотипы; по группе полинезийцев, 10-маркерные гаплотипы, и по группе африканских банту. Идея такая – мы знаем, что цыгане появились в Европе примерно 900-1000 лет назад, в Болгарии – 700 лет назад, поэтому если взять группу их гаплотипов, посчитать мутации, приложить «популяционную» скорость 0.00069 мутаций на маркер на 25 лет, и если получится примерно 700 лет, то скорость 0.00069 – правильная. Разумно, не так ли? Именно это Л. Животовский и объявил, что при калибровке так и получилось, именно со скоростью мутации 0.00069, так и вошло в учебники. То же самое в отношении полинезийцев и банту – у Животовского тоже получилось как надо, скорость 0.00069. А как же, калибровка-то вот она, налицо, так все уже 10 лет и повторяют. Так и ответили авторы той самой статьи, процитированной выше (Hammer et al., 2009), на мои развернутые комментарии в журнале Human Genetics (Klyosov, 2009), в которых я показал, что они считают неверно, ошибаются в 3-4 раза. Авторы сослались на «the original Zhivotovsky et al. (2004) study (i.e., the effective rate was calibrated for time periods of *1,000 years). Since 2004, this approach has been employed by other researchers and critically evaluated in the scientific literature». Перевод: «Исходное исследование Животовского и др. (2004) (содержало) калибровку получающихся скоростей на протяжении примерно 1000 лет». Типа, какие могут быть вопросы, и тем более критика? Скорости калибровались, и всё тут.
Итак, давайте посмотрим для начала на цыган. Гаплотипы цыган были определены у 179 человек из двенадцати болгарских таборов. На удивление, у всех гаплотипы похожи, явно от общего не столь далекого общего предка. Посмотрим сначала, как надо работать с этими гаплотипами, а потом сверим, как «калибровал» Животовский. Для двойной проверки проанализируем как 6-, так и 8-маркерные гаплотипы, и если считаем правильно, результаты должны совпасть, в пределах разумных погрешностей.
У всех 179 цыган гаплотипы в 6-маркерном формате содержали 34 мутации от базового гаплотипа 15-12-22-10-11-12 (в формате DYS 19, 388, 390, 391, 392, 393). Это дает 34/179/0.0088 = 22±4 поколения до общего предка, то есть 550±100 лет. Поскольку из 179 гаплотипов 146 были идентичными друг другу, то есть базовыми, то логарифмический метод дает [ln(179/146)/0.0088] = 23±3 поколениям до общего предка, то есть 575±75 лет назад. Практически полное совпадение результатов по линейному и логарифмическому методу. Это означает, что, действительно, все 179 цыган (или их подавляющее большинство) произошли от одного общего предка. В 8-маркерном формате (дополнительно вошли DYS 389-1 и 389-2) добавилось 25 мутаций, то есть стало 59 мутаций. Получаем: 59/179/0.013 = 25±4 поколения до общего предка, то есть 625±100 лет. В 8-маркерном формате из 179 гаплотипов 126 базовые. Получаем [ln(179/126)/0.013] = 27±4 поколения до общего предка, то есть 675±100 лет назад.
Как видим, все четыре варианта расчета дали расстояние до общего предка болгарских цыган одно и то же, в пределах погрешностей расчета: 550±100, 575±75, 625±100 и 675±100 лет назад. Надо отметить, что хотя присутствие цыган в Болгарии было отмечено 700 лет назад, но эти сведения не обязательно точны, они могут относиться к другим цыганам, не обязательно гаплогруппы Н1 (как в данном случае), потомство у первых цыган могло не выжить, и так далее. Тем не менее, 550-675 лет назад – это не слишком далеко от отмеченного времени «присутствия» цыган в Болгарии.
Читатель, видимо, уже приготовился, что у Животовского получилось значительно больше, то есть не менее 1500 лет до общего предка цыган, и что он повинился, что такого науке не известно, значит, ошибочка вышла с константой 0.00069. Слишком занижена. Ведь цыган было 179, мутаций (в 8-маркерных гаплотипах) 59, так что у Животовского должно получиться 59/179/8/0.00069 = 60 поколений, то есть 1500 лет до общего предка. Но наиболее понимающие читатели, зная популяционных генетиков, уже заулыбались. Потому что у тех раз результат заранее известен, такой и получится. Наука такая. Надо просто уметь рукава пониже держать, желательно, чтобы руки закрывали.
Следим за рукавами. Один гаплотип был из списка удален, потому что он «оказался нечетным» (?). Какой удалять – естественно, на рассмотрение удаляющих (или базовый, или мутированный, то есть направленно вносится искажение серии данных). Далее постулировали, что общий предок всех цыган жил 700 лет, то есть 700/25 = 28 поколений назад. Замечаете прием? Я-то выше рассчитывал, когда жил общий предок (получилось в среднем 600 лет назад), а Животовский просто постулировал, что 700 лет назад, и отсюда вел отсчет. Но есть большая разница – время, когда цыгане были замечены в Болгарии, и время, когда жил общий предок рассматриваемых 179 цыган. Совсем не обязательно, что он был первым цыганом в Болгарии, как уже отмечено выше.
Но 600 и 700 лет – разница не так велика, так как же он подтвердил свою скорость мутации в три раза ниже? (!) А вот как – после совершенно произвольного снятия одного гаплотипа получилось, что при использовании квадратичного метода среднее число мутаций на маркер было 0.01272, и при делении на постулированные 28 поколений до общего предка средняя скорость мутации на маркер оказалась 0.01272/28 = 0.000454. А надо 0.00069, непорядок. Надо данные менять, иначе «теория» под угрозой. Животовский с коллегами написали в статье, что это была «недооценка». Поэтому решили пересчитать данные, введя некие «веса», и в ходе этого сняли еще ряд гаплотипов цыган. После каждой стадии подгонки смотрели, насколько скорость удалось подогнать к 0.00069, но никак не получалось. Тогда решили, что один табор – «Музыканты» – «очень гетерогенный» (на самом деле, по моим расчетам, он был совершенно нормальный, как показано выше при сопоставлении линейных и логарифмических методов), и сняли его целиком, все 19 гаплотипов, и опять ввели «веса по размеру популяции» (что ни в коем случае нельзя делать). Получили среднюю скорость мутации 0.000725±0.000187 (обратим внимание на величину погрешности), и решили, что это и есть 0.00069.
Если это «калибровка», то непонятно, что такое калибровка в понимании авторов «основополагающей» статьи, на которую все попгенетики обязаны ссылаться, иначе рецензенты статью в журнал не пропускают. На это жаловался венгерский автор на форуме по «генетической генеалогии», о чем я уже писал на Переформате. Вот такая популяционная генетика. На самом деле, авторы зря занимались акробатикой, надо было повыкидывать гаплотипы с мутациями как «гетерогенные» или просто дефектные, оставить на оставшиеся полторы сотни человек 23 мутации, и получить 23/150/8/0.00069 = 28 желаемых поколений, то есть как раз искомые 700 лет. «Калибровка» была бы не хуже, и никаких хлопот. Как говорил хрестоматийный герой – «как пожелаем, так и сделаем».
Так же рассчитывали и по гаплотипам полинезийцев (маори, самоанцы и жители островов Кука). Решили исходить из того, что маори прибыли в Новую Зеландию 800 лет назад, то есть 32 поколения назад, по 25 лет на поколение. Гаплотипы были 10-маркерные, но акробатика пошла и там. Самоанцев решили в расчет не включать, а у остальных сократить гаплотипы до 7-маркерных. Видимо, с 10-маркерными не получалось то, что «пожелали». Применяя квадратичный метод, нашли, что среднее число мутаций на маркер в 7-маркерных гаплотипах у маори и островитян Кука равно 0.00998, и разделив на 32 поколения, получили таким образом «форсированную» (в отличие от естественной) скорость мутации 0.00032. Как видно, это вовсе не 0.00069, а в два с лишним раза медленнее. Авторы заключили, что это опять «недооценка», и снова пустились в долгие и вязкие рассуждения, почему это может быть недооценкой. На этом «калибровка» данной серии гаплотипов была завершена.
С африканскими банту и того хуже. Раздел «Материалы и методы». Смотрим про африканских Банту. Читаем – «анализировали 148 человек гаплогруппы Е3а7-М191», перечисляются племена, сообщается, что анализ проводили по 10-маркерным гаплотипам.
Раздел «Результаты». Данных по Банту нет, они вообще в данном разделе не упоминаются.
Раздел «Обсуждение». Банту не упоминаются.
Раздел «Применение». Говорится, что применили полученную величину 0.00069 к популяции Банту, чтобы посмотреть, как это коррелирует с археологическими, лингвистическими и историческими данными. Далее говорится, что если усреднить по всем 11 племенам и популяциям, перечисленным в разделе «Материалы и методы», то получится 3400±1100 лет до «экспансии» Банту в подгруппе со снипом М191. И далее – «археологические и лингвистические данные поддерживают это значение, поскольку показывают присутствие Банту в Западной Африке в неолитические времена, ∼ 1000 лет до нашей эры, или даже 2000 лет до нашей эры». И далее – «Величина 3400 лет назад может рассматриваться как нижняя граница для времен экспансии Банту… Если мы возьмем наиболее частые гаплотипы среди тех 148 хромосом как предковые, и приложим к ним величину 0.00069, то время для М191 получится 14700 лет… В этом случае, экспансия Банту могла произойти ранее, чем 3500 лет назад». И далее идут долгие рассуждения с неопределенной концовкой.
Если это «калибровка», то что такое не калибровка.
К сожалению, исходные данные (гаплотипы) в статье не приведены, ссылка статьи, что они «в сети», не дает никакого линка. Я могу только заключить, что рассматриваемая популяция Банту молодая, не более 1000-1200 лет до общего предка, как это часто бывает с африканскими популяциями, только недавно прошедшими очередное бутылочное горлышко. Ни к каким археологическим или лингвистическим данным это не имеет никакого отношения. Рассуждения в статье не имеют никакого значения в отношении рассматриваемого вопроса. Никакой «калибровки» не было и близко.
Вот такая цена «калибровкам» Л.А. Животовского, которые стали краеугольным камнем популяционной генетики в последние 10 лет. Кроме как «стыд и позор» других слов я не подберу. Последние пять лет я объясняю это в научной литературе, начиная со статьи в Journal of Genetic Genealogy (2009) и Human Genetics (2009), и далее в журнале Advances in Anthropology, выступаю на англоязычных форумах. Но попгенетикам – как о стену горох. Они продолжают плодить в академической литературе мусор. А те, кто в этом ровным счетом ничего не понимают, как тот же воронежский Семененко, тем не менее, вылезают со своим критическим словом:
А профессор Клёсов у них пользуется очень дурной славой. Он считает, что разработал собственную методику определения скорости накопления генетических мутаций, поэтому его «датировки» расходятся с датировками профессиональных генетиков.
Полагаю, смысл этих слов сейчас понятен лучше. Причина «дурной славы» тоже понятна, если почитать выше. Не оставляю я в покое популяционных генетиков с их жульничеством. Правда, одна маленькая деталь. Никакой «дурной славы» в научных публикациях попгенетиков у меня нет. Никто и никогда из них не высказался негативно в отношении моих расчетов. Видимо, Семененко пересказывает приватное, кулуарное жужжание. И на том спасибо, это дало мне возможность рассказать интересующимся, в чем там дело. Поэтому продолжим цитировать Семененко, беря его слова как пример жужжания на тему, в которой он не разбирается. А я продолжу пояснения.
Семененко — Цитирую профессора Клёсова: «Я опираюсь на свои источники, потому что работаю в совершенно новой области науки». У него своя «генетика». А у остальных генетиков мира — своя. Я как-то склонен полагать, что в генетике своих источников, своих методов, своих способов расчётов скорости мутирования генов нет.
Если это действительно моя цитата, то я с ней в целом согласен. Я действительно опираюсь на свои источники, это – кинетика химических и биологических реакций. К генетике она никакого отношения не имеет. Так что фраза Семененко «у него своя генетика» – это бессмыслица. Ошибка – полагать, что направление науки определяется объектом исследования. Изучать ДНК – это далеко не обязательно генетика. Например, химик, растворяя ДНК в кислоте и изучая, скажем, вязкость получаемого раствора, вовсе не занимается генетикой. Альберт Сент-Дьорди как-то сказал – «Дайте химику динамомашину, и он тут же растворит ее в соляной кислоте». Это вовсе не означает, что химик при этом будет заниматься электродинамикой.
Некоторые, не слишком знакомые с предметом, полагают, что ДНК-генеалогия – это популяционная генетика. Это – совершенно не так. Во-первых, популяционная генетика – это часть генетики, о чем говорит ее название, но со своими особенностями. Главная задача популяционной генетики – это выявление связи между генотипом и фенотипом, то, к чему ДНК-генеалогия не имеет отношения. Популяционная генетика зачастую тоже рассматривает гаплогруппы и гаплотипы Y-хромосомы, как и ДНК-генеалогия, но на этом сходство заканчивается. Аппарат популяционной генетики, применяемый для интерпретации получаемых данных, например, «метод главных компонент», характеризуется со стороны ДНК-генеалогии как совершенно примитивный и ведущий к заблуждениям, и не используется в ДНК-генеалогии. Он не отвечает задачам ДНК-генеалогии. Это же относится и к искаженным и примитивным методам обработки картин мутаций в ДНК в рамках популяционной генетики. Если популяционный генетик увидит (или получит) набор из сотни гаплотипов, скажем, 37-маркерных, в котором, например, четыре гаплотипа одинаковых, и еще семь будут совпадать друг с другом случайными парами, он сообщит, что в наборе имеется «89 уникальных гаплотипов». Это и будет результатом его исследования, который пойдет в статью в научный журнал. Ни он, ни рецензенты не хотят признать, это эта «информация» не имеет ни малейшей ценности, и фактически никому в таком виде не нужна. Но таковы принципы и правила популяционной генетики.
«Молекулярной историей» (ДНК-генеалогией) это назвать никак нельзя. А специалист в ДНК-генеалогии сразу скажет, что общий предок этих ста гаплотипов жил 925±105 лет назад, потому что натуральный логарифм отношения 100/4, поделенный на константу скорости мутации для 37-маркерных гаплотипов (0.09 мутаций на гаплотип на условное поколение в 25 лет) равен 36 → 37 условных поколений (стрелка здесь – поправка на возвратные мутации), то есть общий предок этой сотни гаплотипов жил примерно 925 лет назад. Сказать это с большей определенностью можно тогда, когда подсчитано число мутаций во всех ста гаплотипах, и если оно равно 324, тогда 324/100/0.09 = 36 → 37 условных поколений, то есть ровно тот же промежуток времени до общего предка. Погрешность расчетов определяется по известным правилам (Klyosov, 2009). Совпадение времен до общего предка для «логарифмического» и «линейного» метода (в первом мутации не подсчитываются, во втором подсчитываются) свидетельствует, что закономерности образования мутаций в гаплотипах следуют кинетике первого порядка, что, в свою очередь, означает, что все сто гаплотипов действительно произошли от одного прямого общего предка. Популяционная генетика такими подходами не пользуется, они ей чужды по ряду причин, среди них – приверженность устаревшим, традиционным подходам, отсутствие соответствующего образования, клановость, отсутствие хорошей научной школы. Короче, ДНК-генеалогия является другой дисциплиной, нежели популяционная генетика.
Продолжаем цитировать «эксперта» Семененко:
Проблема Клёсова, на мой взгляд в том, что он смешивает научные и ненаучные данные. Т.е. у него есть определённые интересные выводы (связь R1a и R1b с индоевропейцами), но в целом его теорию миграций индоевропейцев я не принимаю, поскольку она явно построена с помощью натяжек и предвзятых данных. Он является ярым инвазионистом относительно истории индоариев в Индии и выводит их из Аркаима, хотя археологически это опровергнуто уже давно. И при этом он сам заявляет, что R1a зародилась в Китае, потом проникла оттуда в Южную Азию, потом исчезла оттуда и вернулась снова в соответствии с канонами инвазионизма/иммиграционизма во II тыс. до н.э. Так что это явный бред с подтасовкой «фактов».
Здесь у Семененко – гроздь недоразумений. Видимо, «критик» не в ладах с разницей понятий «данные» и «интерпретации». Какие, интересно, «ненаучные данные» я с чем-то смешиваю? Что это за «ненаучные данные»? Гаплотипы? Гаплогруппы? Субклады? Снипы? И что же там «ненаучного»? Если это «теория миграций индоевропейцев» – то это уже не данные, а интерпретации. Видимо, Семененко не понимает, что «теория» и «данные», это, так сказать, две большие разницы. Я вполне допускаю, что некоторые мои интерпретации допускают другую трактовку, в них возможны подвижки, которые будут появляться с появлением нового знания. Но если Семененко «не принимает», как он пишет, то это не аргумент, это нечто низкопробное. Как и «построена с помощью натяжек и предвзятых данных». Какие там натяжки и предвзятые данные – Семененко, разумеется, не говорит. Это в моей классификации – «ля-ля». Для тех, у кого нет упомянутых ранее рецепторов – это любимое дело. Сотрясать воздух, не приводя никаких предметных аргументов. Выйдите, покажите, где там «натяжки» и «предвзятые данные», предложите свои объяснения, непременно основанные на фактах, на экспериментальных данных, тогда и поговорим.
Что касается «связи R1b с индоевропейцами», то там Семененко опять что-то не понял, и в любом случае изложил невразумительно. Не было у R1b никакого «индоевропейства» до начала I тыс. или конца II тыс. до н.э. Нигде, где они проходили, индоевропейских языков не оставили – ни на севере Казахстана, ни на Средней Волге 8-6 тыс. лет назад, ни на Северном Кавказе 7-6 тыс. лет назад (там они оставили другие языки), ни в Месопотамии, ни у шумеров, ни в Египте, ни у басков. Ни потом в Европе до конца II тыс. до н.э. Везде там были (или кое-где остались) другие, неиндоевропейские языки. Индоевропейские языки появились у эрбинов (носителей гаплогруппы R1b) только во времена кельтов, в первой половине I тыс. до н.э. Это кельты, судя по данным ДНК-генеалогии, принесли ИЕ языки в Европу, причем, видимо, с гаплогруппой R1a. Сейчас, разумеется, подавляющее большинство носителей гаплогруппы R1b в Европе говорят на ИЕ языках, но попгенетики всегда переносят то, что есть сейчас, на то, что было тысячелетия назад. Поэтому в их понимании R1b всегда говорили на индоевропейских языках, раз говорят на них сейчас.
Но давайте разберем цитату «эксперта» по частям. Часть первая: Он является ярым инвазионистом относительно истории индоариев в Индии и выводит их из Аркаима, хотя археологически это опровергнуто уже давно.
Я был бы признателен за сведения о том, что там «археологически опровергнуто». Что именно? Что в Аркаиме были не арии? Что они не прошли в Индию примерно в середине II тыс. до н.э., а, возможно, еще и ранее, и это «опровергнуто археологически»? Я не знаю, что такое «инвазионист», и тем более «ярый», но догадываюсь, как в том старом анекдоте известное радио не знало, что такое горжетка, но догадывалось. Семененко вообще как-то вязко пишет, что-то у него с изложением мыслей не так. Что такое «выводит их (видимо, индоариев) из Аркаима»? На самом деле Аркаим – это транзитный путь ариев, изначально выводить оттуда их просто не получается. Или они там завелись сами по себе, как мыши в грязном белье (была такая теория)? Или Семененко полагает, что в Аркаиме были не «индоарии», а «иранцы» (был такой спор среди археологов), но я эти термины не использую (хотя не мешаю использовать другим), назвать их ариями будет достаточно.
Л.С. Клейн в своей недавней книге «Этногенез и археология» (том 2, стр. 39) пишет об обнаружении в Южном Приуралье (Синташта) погребений с конями, колесницами, изображением свастики. Или это не арии? Что там «отвергнуто археологически»? Видите, какой это дефект, любезный А. Семененко, когда критик не может связно выражать мысли. Приходится гадать, что он намеревался сказать, потому что в итоге получилась некая каша.
Читатель задает ему вопрос: «Александр, а Вы бы не могли ответить, пожалуйста, какие именно работы современных исследователей в области молекулярной генеалогии Вы использовали в своих работах? И насколько результаты этих исследований совпадают с данными других наук? Насколько точны данные ДНК-генеалогии? Есть ли разночтения в датировках (этногенез, миграции)?»
Семененко, как для него характерно, на прямой вопрос не отвечает. Ни на то, какие работы современных исследователей он использует, ни на то, как эти данные соотносятся с данными других наук, ни на вопрос о точности данных ДНК-генеалогии (этого он уж точно не знает), ни на вопрос о датировках. Вместо этого он приводит список своих статей, все местных региональных конференций. Не густо, пожалуй. Посмотрим, что там.
Первый источник – типичная провинциальная компиляция: кто, что, когда, о чем сказал. Иванов сказал то-то, Петров ему возразил, Сидоров высказался иначе. Шлегель указал, Лэтэм отметил, Мюллер указал, Мюир отметил, Уилсон поправил, Якоби, Бюллер, Винтернитц, Вулнер, Грисволд, Моултон, Калла, Норман, Маршалл, Уилер, опять Мюллер, Казанас, и все что-то отметили или указали. Своего, от Семененко, ровным счетом ничего нет, ни единого своего слова, никаких своих данных. Вплоть до конца статьи – сплошные ссылки на других. Вывод: «как говорит ссылка 52» (набор авторов из разных книг и статей – Шаффер и Лиштенштейн; Эрдоси; Витцель; Кенойер; Элст; Кордона и Джейн; Мак-Интош), археологических следов арии во II тыс. до н.э. не оставили, а «как говорит ссылка 53» (опять десяток книг и статей, сообщение, что древнеиндийские тексты «об этом» (об отсутствии археологических следов?) не говорят. О чем «об этом»-то? Как можно быть таким косноязычным? А «как говорит ссылка 54», между 6500 и 2800 лет назад, как было найдено, имеется непрерывный ряд однотипных костных останков человека, то есть не было «ощутимых вливаний извне». Ссылки, правда, все 15-20-летней давности, или вообще без указания года. Наконец, «как говорит ссылка 55» (одна работа 77-летней давности, еще три без указания года издания, в том числе под названиями «Лингвистические аспекты», «Ригведная Индия» и тому подобное), «индоарии не могли прийти в Индию после 4500 лет до н.э.», то есть позднее 6500 лет назад. Всё, конец статьи. Статья ссылкой «как говорит» и закончилась.
Доказательств – по сути никаких, все столь же вязко. В качестве доводов (хотя их и доводами назвать нельзя) – те же перепевы про Митанни, про индоариев, кто куда ходил – ничего нового, все это повторяется десятилетиями, компилируется, пересказывается, повторяется. Читать откровенно неинтересно. Неинтересно еще и потому, что ДНК-генеалогия в корне противоречит этим соображениям, потому что большинство носителей гаплогруппы R1a в Индии имеют субклад R1a-L657, который образовался всего примерно 4000 лет назад, и шлейф его уходит в Индию со стороны Европы. В Европе его очень мало, в Индии – миллионы человек. Это – те самые «индоевропейцы», потомки древних ариев. Датировки ДНК-генеалогии стыкуются с временами прихода ариев в Индию во II тыс. до н.э., причем именно с севера или северо-запада. В общем, это можно обсуждать, но не с Семененко. Что же касается археологии, то советую Семененко почитать хотя бы последнюю книгу Л.С. Клейна «Этногенез и археология», глава «Откуда арии пришли в Индию». Хоть я Клейна и ругаю за непонимание и за нежелание понять ДНК-генеалогию, но археологический материал он собирает довольно аккуратно.
Так вот, упомяну просто на ходу, что археологи прослеживают миграцию ариев из Причерноморья (катакомбная культура) в Индию, и это неплохо доказано еще с 1970-х годов, то есть уже больше 40 лет. Что-то я у «индолога» Семененко я этого не видел. А катакомбная культурная общность располагалась к западу от срубно-андроновского круга в первой половине II тыс. до н.э., то есть примерно 4000-3500 лет назад. И далее, как отмечает Клейн, цепь катакомбных памятников и культур выявлена между Северным Причерноморьем и Индией, причем их возраст уменьшается по мере продвижения от Причерноморья к Индостану. И далее Л. Клейн подробно описывает материальные признаки из погребений, которые свойственны как ариям Причерноморья и по всему указанному выше пути, так и ариям в Индии. Так что советую Семененко учить материальную часть.
А что до того, что археологи не смогли найти в костных останках ариев, прибывших в Индию во II тыс. до н.э., точнее, так интерпретируют свои данные, то есть хорошее правило – «отсутствие доказательств не есть доказательство отсутствия». Еще один совет Семененко – не переписывать тупо то, что написали другие, а заняться делом самому: поднять первичные данные археологов (а не их мнения), и внимательно пересмотреть их под углом того, что арии в Индию приходили, во II тыс. до н.э., как и свидетельствует ДНК-генеалогия. Конечно, картина могла быть более сложной, поскольку ДНК-генеалогия пока рассматривает ДНК ныне живущих индийцев, и «индоевропейские» индийцы (их так и классифицируют в Индии) гаплогруппы R1a в своем большинстве имеют общих предков – по разным сериям гаплотипов – живших между 4000 и 3500 лет назад. 4000 лет назад – это, похоже, еще не Индия, а Русская равнина, Средняя Азия или Южный Урал, в общем, северные территории.
Дальше читать его сочинения расхотелось по причине откровенной некомпетентности А. Семененко, но я сделал усилие и открыл следующее, под малообещающим названием «Вклад антропологии и генетики в решение индоарийской проблемы», опять региональная научная конференция. Малообещающее – потому что если вклад есть, то так и надо статью назвать, по конкретному вкладу. А статьи под названиями типа «Вклад…» или «К вопросу о…» читать как-то не хочется. Ясно, что там мало что есть. Так в данном случае и оказалось. Статья – просто кошмар. Во-первых, вся подчистую переписана из статей популяционных генетиков, причем именно тех, кто применяли ту самую печально известную «популяционную скорость Животовского». Во-вторых, как следствие во-первых, в статью Семененко аккуратно перенес все те чудовищные ошибки попгенетиков, а именно завышения датировок прихода гаплогруппы R1a в Индию на 250-350%. Отсюда у Семененко и оказался в статье просто кошмар. Безграмотный анализ попгенетиками картины и хронологии мутаций в ДНК, и безграмотное перенесение всего этого Семененко в свою статью дало очередной мусор. Слава Богу, дальше Воронежа, видимо, это не пошло.
Итак, что у него (точнее, у попгенетиков, с которых он срисовал ошибочные данные и ошибочные трактовки) получилось? А получились сапоги всмятку. Если цитировать этот «кошмар», то нужно всю статью цитировать, от начала до конца. Поскольку датировки по Индии были завышены в несколько раз, то якобы получилось, что датировки тех, чьи предки жили 3500-4000 лет назад, уехали в глубь времен до… В общем, предоставим слово Семененко, который любит писать заумно: « Эпоха аккумулированной микросопутствующей вариативности в большинстве индийских гаплогрупп превышает 10000-15000 лет, что соответствовало древности региональной диференциации». Вспоминается – «Аркадий, не говори красиво». А поскольку попгенетики завысили датировки гаплогруппы R1a по Индии (от 3500-4000 лет до 10000-15000 лет, то есть, как обычно, на 250-400%), а по Европе не считали, и взяли обычные 4500-5000 лет, то, как старательно цитирует Семененко, «оказалось, что индийцы отражают более высокое разнообразие в гаплогрупппе R1a1 даже при сопоставлении с южно- и восточно-европейским населением» («разнообразие» – это эвфемизм попгенетиков, когда они датировки рассчитать не умеют, но сказать хочется). Отсюда, естественно, следующий безграмотный вывод – «Неизбежным становился вывод о том, что индоевропейцы возникли в Индии и оттуда распространились в Европу» (ссылка на попгенетиков, фамилии те же, что были упомянуты выше, в данном случае эстонец Кивисилд с коллегами, 2002-2003 гг.). Эстонская «школа» попгенетиков – еще та. Во всяком случае, в отношении датировок и последующих безумных трактовок исторических событий.
Для полноты дискуссии следует отметить, что европейские R1a на много уровней субкладов древнее «индоевропейских» субкладов R1a в Индии. Вот – сокращенная схема:
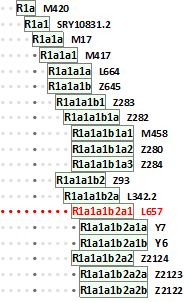
Индийские характерные «индоевропейские» субклады, потомки арийских, начинаются только с красного уровня, это R1a-L657. Они – «внуки» европейского субклада R1a-Z93, который образовался в Европе примерно 5200 лет назад; его ближайший потомок, L342.2 – примерно 4200 лет назад; «внук», L657 – примерно 3500 лет назад. Из него никак не могли произойти все те европейские (и, возможно, азиатские) субклады, которые находятся выше красной линии. Так что преобладающий в Индии субклад L657 никак не мог породить европейские R1a линии. Попгенетики, которых обильно цитирует Семененко, этого не знали, с датировками чудовищно ошиблись, но в глазах Семененко – они – представители «международных коллективов профессиональных генетиков, опубликованные в ведущих международных научных журналах по генетике». Вот так некомпетентность и слепая вера в «авторитеты», которая в науке совершенно неуместна, приводит к серьезным конфузам.
В связи с этим забавно читать у Семененко, что «генетики использовали высокоточные данные… по 10 микромаркерам». Для начала, микромаркеров не бывает. Маркер – он и в Индии маркер. Генетики используют порой термин «микросателлиты», здесь Семененко в порыве рвения копирования у генетиков перестарался. Но занятно здесь понятие «высокоточные», в применении к 10-маркерным гаплотипам. Не случайно Семененко так и не ответил на вопрос читателя «Насколько точны данные ДНК-генеалогии?» В этом он, ясно, тоже не разбирается.
Далее, это в далеких 2002-2003 гг. характеристика гаплогруппы R1a ограничивалась снипом R1a-М17, что и цитирует Семененко. Сейчас мы уже знаем об этой гаплогруппе в Индии и вокруг значительно больше, оперируем субкладами Z93, L342.2, L657, Z2124, Z2123, Z2122 и другими, в целом, знаем времена их возникновения и направления миграций. Знаем, что в отдаленных провинциях в Индии, в племенах (которые в касты не входят) живут носители более древних ветвей гаплогруппы R1a, но это не индоевропейские группы, и они пришли, видимо, с востока, возможно, из Алтайского региона более 12-15 тысяч лет назад. Но указанные попгенетики с этими гаплотипами не работали, у них бы получились датировки более 40 тысяч лет назад. Они, эти древние «автохтоны», из Индии в последние 10 тыс. лет никуда не выходили. Этим вопросом нужно специально заниматься, но данных по ним очень мало, субклады неизвестны. Они, эти люди индийских джунглей, свои образцы ДНК в американские лаборатории с приложением 200 долларов не высылают. Да сейчас разговор и не о них.
Не понимая сути ДНК-генеалогии, смешав все и вся в отношении датировок, гаплотипов и древних миграций, Семененко пишет свою статью как плохой анекдот. То, что носители гаплогруппы I не ушли с ариями в Индию он тоже засчитывает в то, что арии в Индию не приходили. Иначе, по его мнению (!), обязаны были прийти, ведь «предполагаемый возраст гаплогруппы I – 22000 лет, этого возраста достаточно для экспансии, и она также намного чаще встречается в Европе по сравнению с гаплогруппой R1a1». Здесь все перепутано, не понято и искажено. Дело в том, что древняя гаплогруппа I практически исчезла в Европе, видимо, в IV-III тыс. до н.э., и возродилась (из немногих выживших потомков) только примерно 3600 лет назад в виде гаплогруппы I1, и примерно 2300 лет назад в виде гаплогруппы I2 на Дунае. Они просто не могли уйти с ариями 4500 лет назад, они, эти гаплогруппы, мучительно выживали в те времена, с их носителями в минимальных количествах. Так что никакой экспансии не было до конца прошлой эры, когда гаплогруппа I2 размножилась на Балканах, и это только там их численность сейчас превышает (и то не везде) численность гаплогруппы R1a.
И вот такими пассажами заполнена статья Семененко. Там же – подобные рассуждения про гаплогруппу «N3», чей индекс давно устарел. Семененко, переняв у попгенетиков манеру рассматривать современные «частоты» гаплогрупп и автоматически перенося их на тысячелетия назад, опять создает совершенно искаженную картину их взаимоотношений с ариями, рассматривая свои (неверные) представления о «миграциях R1a1 c востока на Балканы» в арийские времена. Опять он, Семененко, пространно цитирует статью Underhill et al. (2010), в которой опять использовали «популяционные скорости мутаций» Животовского, и получили, разумеется, датировки в мезолите, когда там на самом деле времена 3500-4000 лет назад. В итоге, окончательно запутавшись, Семененко провозглашает, что данные попгенетиков (в его пересказе) «исключают Европу из списка прародины индоариев», и что их «миграции начались не в Европе». Здесь можно было бы начать обсуждение, что такое «прародина» в данном контексте, и что такое «миграции начались», но, конечно, не с Семененко, при его фантастической некомпетентности в данном вопросе. Понятно, что он хочет сказать, и на чем основано это его хотение, но это продолжения дискуссии не стоит.
Остальные его опусы я уже не читал. Нет смысла.
Поехали дальше, часть вторая, и последняя:
И при этом он сам заявляет, что R1a зародилась в Китае, потом проникла оттуда в Южную Азию, потом исчезла оттуда и вернулась снова в соответствии с канонами инвазионизма/иммиграционизма во II тыс. до н.э. Так что это явный бред с подтасовкой «фактов».
Опять искажения. Во-первых, не «зародилась в Китае», Китая тогда не было. Не зародилась, а обнаружена на севере современного Китая в количествах до 30% от нескольких этнических групп. Туда она, скорее всего, попала из алтайского региона, из Южной Сибири. И ничего удивительного в этом уже нет, как это представлялось ранее, так как именно в Сибири недавно найдены ископаемые гаплогруппы K и R. Гаплогруппа R мигрировала (в своей части) в Европу в виде дочерних гаплогрупп R1a и R1b, гаплогруппа R1b так в Европе и осталась с начала III тыс. до н.э., то есть со времени образования культуры колоколовидных кубков примерно 4800 лет назад, а гаплогруппа R1a, будучи, видимо, вытесненной из Европы (как и многие другие гаплогруппы), перешла на Русскую равнину и разошлась по разным направлениям как исторические арии – на юг, в Месопотамию и Митанни, и далее в Ирак, Саудовскую Аравию и страны Персидского залива; на юго-восток, в Среднюю Азию и далее на Иранское плато; на восток и затем на юг, в Индию; и далее на восток, в Зауралье, в скифский круг археологических культур. Никакой «подтасовки фактов» здесь нет и близко. Датировка ископаемой R1a на территории современной Германии (Эулау) – 4600 лет назад, датировка R1a в Зауралье, андроновская культура, 3800-3400 лет назад. При чем здесь «инвазионизм», который у Семененко исполняет роль жупела? А далее на Алтае – пазырыкская культура, скифский круг, опять R1a, датировка – начало I тыс. до н.э., и далее, вплоть до начала нашей эры. Можно поподробнее про «подтасовку фактов»?
Думаю, подобный «разбор полетов» полезен для тех, кто хочет лучше разобраться в ДНК-генеалогии, кто хочет понять воистину пагубную роль «популяционной генетики» в отравлении мозгов, как оказались отравлены мозги Семененко (не буду иронизировать глубже), который без критического анализа бросился с объятиями к популяционной генетике и ее принципиально искаженными подходами и соответствующими выводами. Что касается персональных выпадов в мой адрес – мне они, по сути, безразличны. Я их использую для преподавания подходов и принципов ДНК-генеалогии, для изучения все увеличивающейся роли ДНК-генеалогии в исторических науках и языкознании. Но интересная закономерность – как только я рассматриваю подобную «критику» – тут же вылезает впечатляющая некомпетентность «критиков», их неспособность мыслить и сопоставлять факты, наблюдения, интерпретации, закономерности. И при этом – неуемное желание вылезти на публику и обнародовать их «критику». Заметьте – не попытаться вынести что-то для себя полезное, нет, а именно отвергнуть, не пытаясь вдуматься. Здесь определенно есть что-то системное. Но это вопрос уже к психиатрам, не ко мне.
Анатолий А. Клёсов
доктор химических наук, профессор
Перейти к авторской колонке
| Сообщение от Переформата: серьезная просьба интересующимся – направить короткое сообщение с предварительной заявкой на тест. Если не будет первой тысячи участников, то проект московской ДНК-лаборатории притормозится до лучших времен. Спасибо тем, кто уже написал, таких уже немало! Повторно просим не отправлять, всё что вы отправили – было принято. Заявку можно оставить здесь, никаких финансовых обязательств она не несет, просто сообщение о намерениях, если вас в будущем всё устроит. |

















