
Почти год назад на Переформате вышла моя статья «Суета попгенетиков вокруг генома». Статья, как говорит хрестоматийное высказывание, резкая, но справедливая. В ней нередко повторялось слово «акробатика», и с самого начала было отмечено – «в основе этой акробатики – опять, увы, «популяционная генетика человека» в своем наихудшем виде. К другим положениям той статьи мы опять сегодня вернемся, поскольку в полном соответствии с законами диалектики ситуация с суетой попгенетиков вокруг генома развивается по спирали, переходя от плохой к более худшей.

В чем проблема, и зачем ее здесь обсуждать, в круге читателей, казалось бы, довольно далеких от академических изысканий попгенетиков? А в том, что проблема намного шире, чем академические изыскания. Она в том, куда ведут келейные исследования замкнутых научных коллективов, отгородившихся корпоративным забором своих рецензентов и редакторов изданий, коллективов, не имеющих добротной и критической (что во многом одно и то же) научной школы, которые не хотят (возможно, и не могут) осваивать другие подходы, хотя бы для того, чтобы независимо проверять свои, обычно примитивные и, по сути, часто неверные построения. Как я не раз саркастически объяснял, основный принцип работы попгенетиков – «что вижу, то и пою». Исторического осмысления просто нет. Его же не видно, там думать надо. Критически размышлять.
И вот появилось новое поле, в своем нынешнем развитии – появилось буквально в последние год-два. Поле для «что вижу, то и пою» – бесконечное. Это – геном человека. И вот теперь до него дорвались популяционные генетики. Что это поле показывает? А показывает оно сотни тысяч и миллионы необратимых (или практически необратимых) мутаций (так называемых снип-мутаций, от сокращения SNP – Single Nucleotide Polymorphism) в ДНК самых разных людей планеты. Дело за малым, а компьютер все стерпит.
Суть методологии попгенетиков – сопоставлять эти самые сотни тысяч и миллионы мутаций, и находить между ними общности. Эти мутации закладываются в компьютер, и компьютер показывает, что вот у того австралийского аборигена, для которого выявлено 423 567 мутаций, и этого американского индейца, у которого найдено 502 017 мутаций, у них 26 789 мутаций общие, одни и те же. Значит, ясно, что американский индеец произошел от австралийского аборигена. Вот, смотрите, компьютерная картинка, построена по специальной программе, вот их общие мутации, на диаграмме раскрашены одним цветом, какие еще вопросы? Как, вы спрашиваете, а почему тогда не наоборот? Что это абориген произошел от индейца с таким же успехом? Ну как же, как же, мы же знаем, что Америку заселили только 16 тысяч лет назад, а Австралию – 50 тысяч лет назад. Не может быть наоборот.
Значит, записали – выявили наследственные корни американских индейцев, крупный успех современной науки, впервые в мире и так далее, вот и статья готова в ведущее академическое издание. Там и тысячи мутаций описаны, и методология секвенирования ДНК, и компьютерные программы приведены, и цветные картинки-диаграммы, и личное согласие индейца и австралийского аборигена процитировано о том, что они согласны на изучение их генома, в соответствии с Хельсинкской декларацией, особым положением об этике научных исследований. Статья как по маслу проходит в ведущее академическое издание, рецензенты все свои, никто не читает и не разбирается, более того, их сам автор теперь и предлагает – зачем здесь сюрпризы, дело денежное, поскольку потом на основании этой статьи получаются финансовые гранты – и на зарплату, и на приборы и оборудование, и на сбор материала у очередных обитателей острова Пасхи, и, само собой, для поездки на этот самый остров. Статья выходит, пресса в восторге сообщает о Breaking News, сенсация, ведущие журналы и газеты об этом пишут как об очередном эпохальном открытии, тысячи линков в сети, бурные обсуждения, все пляшут и поют.
И нет ни одного здравого человека в этой цепи абсурда, который бы сказал – минуточку, а ведь эти выводы, что индеец от австралийца произошел – ерунда на постном масле. Потому что, смотрите – американский индеец относится к Y-хромосомной гаплогруппе Q, а австралийский абориген – к гаплогруппе С. У них есть общие предки – и гаплогруппа ВТ, и гаплогруппа СТ, и гаплогруппа CF, как показывает дерево гаплогрупп ниже. Не говоря об общем предке человека и шимпанзе, от которого и у индейца, и у австралийца масса общих мутаций. Но мутации от шимпанзе обычно «отфильтровывают», так что их можно во внимание не принимать. А остальные снип-мутации остались и у индейца, и у австралийца. И вот вы на них и смотрите. Они, эти мутации показывают, что ни один от другого произошел, а оба произошли от общего предка. Так что сенсация – дутая, как и выводы статьи, и все остальное.

Объяснение крайне простое, но по какой-то загадочной причине попгенетики до него не доходят, и продолжают гнать статьи о том, кто от кого сенсационно произошел, наводняя ведущие академические журналы статьями такого рода. Иначе говоря, они сопоставляют снип-мутации линейно, одномерно, а надо смотреть на общих предков в системе 3D. Рецензенты этих статей тоже про это или не знают, или статьи, им присланные на рецензию, просто не читают. Они смотрят на фамилии авторов по системе свой-чужой. Раз свой – то проходи, пропуск можно не показывать. А то в следующий раз и мою, рецензента, статью могут и прочитать, а сюрпризы нам опять не нужны. Вот такая за последние годы сложилась система в академических журналах.
На самом деле я эту ситуацию с австралийским аборигеном и американским индейцем не придумал. Только что вышла статья (опубликована 30 декабря 2013 года; если кто хочет ее найти и почитать на досуге – A.M. Ribeiro-dos-Santos и др. [всего 12 авторов], High-Throughput Sequencing of a South American Amerindian, PLOS One, т. 8, № 12), в которой вот так же формально обрабатывали 502 017 снип-мутаций по всему геному (точнее, анализировали 95.92% генома) одного южно-американского индейца гаплогруппы Q1a3a. Авторы заключили, что хотя они продолжают придерживаться широко принятой концепции прибытия будущих американских индейцев из Сибири через Берингию, полученные ими данные по геному «принесли новые сведения для понимания истории популяций человека».
Если кто еще пока не очень уловил, в чем абсурд многих исследований популяционных генетиков по «линейному» сопоставлению геномов, приведу несколько примеров восходящей сложности, в которых иллюстрируется, к чему приводит такой одномерный подход.
Пример 1. Изучаются геномы родственников, мальчика и его дяди (брата отца). Геномы в большой степени перекрываются, как и должно быть. Полному перекрыванию мешает то, что мама мальчика (жена отца мальчика) имеет свой геном, поэтому геном ее сына «разбавлен» по сравнению с геномом отца мальчика. Ну, а геномы отца мальчика и его брата, дяди мальчика, практически идентичны.
Что делает попгенетик? Он видит, что геномы мальчика и дяди перекрываются, и говорит: это показывает, что дядя – предок мальчика. Понимаете, что такое линейное мышление? Это когда не смотрят в глубь явлений. А на самом деле нетрудно понять, что предок обоих – дедушка. Даже если не знать, что это дядя и племянник. Можно ведь понять, что перекрывание геномов не говорит о непременной прямой наследственности, а говорит только о том, что у них – общий предок. Когда он жил – показывает дополнительный анализ, и для этого есть достаточно налаженные методы, например, методы ДНК-генеалогии. Тогда достаточно посмотреть на гаплотипы обоих тестируемых, увидеть разницу между ними в числе мутаций, и определить, когда жил их общий предок. Другими словами, тогда мы будем иметь и геномную картину обоих, и гаплотипы обоих, и расчеты хронологии до их общего предка, чего геномная картина обычно не позволяет или получает это с огромной погрешностью. Как, например, если с помощью микроскопа пытаться изучать звездное небо.
Просто и наглядно, не так ли? Нужно сочетать геномные исследования с ДНК-генеалогией, и получать более полную и более правильную картину истории популяций. Но попгенетики этого не делают, что будет показано в этом очерке. Их ДНК-генеалогия пугает, они ее не понимают, и никак между собой не договорятся, как к ней подходить. Все это давно опубликовано, но попгенетики читают только статьи, написанные попгенетиками, оставаясь за тем забором, что обрисован выше.
Пример 2. Сравниваются геномы двух мужчин, назовем их Александр Иванович и Анатолий Алексеевич. С протяженными гаплотипами попгенетики не работают, но, допустим, им показали и они поняли, что им показывают 67-маркерные гаплотипы этих двух мужчин. Между ними – четыре мутации.
13 24 16 11 11 15 12 12 10 13 11 30 – 16 9 10 11 11 24 14 20 34 15 15 16 16 – 11 11 19 23 15 16 17 20 37 41 12 11 – 11 9 17 17 8 11 10 8 10 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13
13 24 16 11 11 15 12 12 10 13 11 30 – 16 9 10 11 11 24 14 20 34 15 15 16 16 – 11 11 19 23 15 16 17 21 36 41 12 11 – 11 9 17 17 8 11 10 8 10 10 12 22 22 15 10 12 12 13 8 15 23 21 12 13 11 13 11 11 12 13
Если кто читал очерк про детей боярских, тот знает, чьи это гаплотипы. Знает уже, что четыре мутации между ними показывают, что общий предок этих двух людей жил примерно в 1588 году, и знает, как это было рассчитано. Попгенетики этого не знают, и учиться не хотят. Константы скоростей мутаций в 67-маркерных гаплотипах они тоже не знают и не понимают, это многократно проверено. Они все еще считают, что это науке неизвестно.
Так вот, как только попгенетик увидит эти два гаплотипа выше, первое, что он сделает, это разделит 4 мутации на 67, и найдет, что мутации составляют 5.97% от числа маркеров в гаплотипе. Зачем это нужно – он не знает, и на самом деле это никому ни для чего не нужно. Как никому не нужно и «генетическое разнообразие» гаплотипов, которое попгенетик непременно рассчитывает. Никто никогда и ничего путного из этого «генетического разнообразия» не получал. Но если можно рассчитать и поместить в статью – то почему нет? А в ДНК-генеалогии нет «разнообразия» как показателя, есть несравненно более конкретная величина – когда жил общий предок рассматриваемой серии гаплотипов, сколько лет назад. Это – не абстрактная, ничего не значащая цифра, а исторически значащая величина.
Так вот, попгенетик найдет, что геномы Александра Ивановича и Анатолия Алексеевича перекрываются, и запишет, что один произошел от другого. Кто от кого произошел – это попгенетик придумает, возьмет с потолка. То, что геномы перекрываются, потому что оба произошли от одного общего предка – это попгенетику в голову не придет.
Некто спросит – неужели они, в самом деле, такие… в общем, тупые? Трудно поверить, что настолько. И мне трудно поверить, я бы хотел найти другое объяснение, но пока не нашел. Я сам страдаю, когда читаю их «исследования». Если кто думает, что я преувеличиваю, посмотрите на следующие примеры.
Пример 3. Попгенетики нашли, что в геномах русских в среднем 11.7% генома американских индейцев. Стало быть, америнды – предки русских. И не только русских, а и ирландцев (около 8% американских индейцев), украинцев (8.5% америндов), поляков (8.2% америндов), литовцев (9.1% америндов). Есть уже серия статей по этому вопросу. Сенсация, не так ли? Но откуда попгенетики это взяли?
А все оттуда же – геномы частично перекрываются. Но попгенетик понимает, что русские – вряд ли предки американских индейцев. Значит, наоборот. Почему, как это могло быть – попгенетика не интересует. Это уже не его вопрос. Его вопрос – записать, доложить, написать статью, вызвать интерес масс-медиа, получить следующий грант.
Но если мы посмотрим на дерево гаплогрупп выше, мы сразу увидим, в чем там дело. Вот – фрагмент этого дерева, несколько расширенный:

Американские индейцы – в основном гаплогруппа Q, русские, украинцы, поляки, литовцы – в основном гаплогруппа R1a, ирландцы – в основном гаплогруппа R1b. Они все, в основном, происходят от общего предка – гаплогруппы Р, как и индейцы гаплогруппы Q.
Поэтому геномы, ясное дело, перекрываются. Точно так же, как и в примере про мальчика и его дядю. Ну что, трудно было попгенетику про это сообразить? А ведь ни в одной статье о происхождении русских и прочих ирландцев от американских индейцев гаплогруппы даже не упоминались. А зачем это попгенетику? Он же ведь с геномами работает. Что видит, то и поёт.
Пример 4. Совсем недавно, в октябре 2013 года в знаменитом журнале Science была опубликована статья о том, что на верхнепалеолитической стоянке Мальта, в 85 км к западу от Иркутска, в костях четырехлетнего мальчика с датировкой 24 тысячи лет назад была выявлена гаплогруппа R. Мировая сенсация. Газеты запестрели интервью с ведущими популяционными генетиками, антропологами, историками, которые выражали полный восторг от того, что европейская гаплогруппа R оказалась в Южной Сибири, да еще в столь давние времена. Один крупный попгенетик сказал, что это надо переварить, осознать гигантскую значимость этой находки. Похоже, что они не читают Переформат.
Еще в марте прошлого года в статье «Суета попгенетиков вокруг генома» я писал: «…О том, что они (R1b) и в Сибири есть – авторы во внимание не принимают… Популяционная генетика». За год до того, в апреле 2012 года, вышла моя большая статья в журнале Advances in Anthropology («Успехи Антропологии»), которая называлась «Древняя история эрбинов, носителей гаплогруппы R1b, из Центральной Азии в Европу, от 16 тысяч лет назад…», где по всей статье повторяется, что гаплогруппа R1b образовалась в Южной Сибири, естественно, от гаплогруппы R1, и давно опубликованы мои расчеты, что гаплогруппа R1 образовалась, естественно, из гаплогруппы R там же, в Южной Сибири, 26 тысяч лет назад. А та образовалась там же из гаплогруппы Р, которая прибыла в Южную Сибирь примерно 40 тысяч лет назад.
Ну, это ладно, попгенетики статей не читают, об этом уже неоднократно говорилось, и беспокоиться мне за них в этом отношении нечего. Не об этом (точнее, не только об этом) мой сарказм, а о том, что, как водится, попгенетики в связи с этой находкой гаплогруппы R погнали волну абсурда. Они определили геном по костным останкам того мальчика с Байкала, и нашли, что он перекрывается с геномом американских индейцев.
Некоторые читатели моего очерка, наверное, уже заулыбались, потому что запомнили ту небольшую диаграмму чуть выше, на которой гаплогруппы R (мальчика с Байкала) и Q (американских индейцев) сидят рядом, и, естественно, их геномы обязаны перекрываться, поскольку они происходят от одного общего предка – гаплогруппы Р. Они и перекрываются – на величину 14-38%. Это – совершенно нормальное дело, и ровным счетом ничего не означает, кроме того, что перед нами – родственные гаплогруппы.
Но у попгенетиков – свой абсурдный мир. Они восприняли это перекрывание геномов как знак, что люди с Байкала гаплогруппы R шли в Америку, в итоге пришли, и принесли свой геном американским индейцам. А шли они, естественно, из Европы, поскольку сейчас гаплогруппа R, точнее, дочерняя ей R1b – основная западноевропейская гаплогруппа. Вот и найдена история предков американских индейцев! Опять сенсация! Опять пошла горячка «акул пера и шакалов ротационных машин», как отмечали хрестоматийные классики советского юмора и сатиры.
Смех-смехом, а дело было серьезное. Вот заголовок статьи в Science в конце октября – «Ancient DNA links Native Americans with Europe», то есть «Древняя ДНК связывает американских индейцев с Европой». Заголовок статьи в журнале Nature в конце ноября – «Верхнепалеолитический сибирский геном показывает двойное происхождение американских индейцев». Ведущий комментатор Диенекес Понтикос, который вызывающе не имеет понятия о ДНК-генеалогии (о чем я не раз писал – и в своей книге «Происхождение славян», и в его блоге) – «самое интересное, что это показывает поток генов между останками на Байкале и американскими индейцами… похоже, что поток генов был именно от МА-1 (так окрестили в западной прессе древнего мальчика из Мальты) к американским индейцам, а не наоборот». И не понимает Понтикос, что никакого «потока генов» к американским индейцам не было ни от мальчика, ни от его родителей, ни потомков его братьев. Равно как не было и «двойного происхождения американских индейцев». Поток генов был от их предка, из гаплогруппы Р, жившего примерно 15-20 тысяч лет до того, в гаплогруппу Q, и уже они ушли в Америку. А носители гаплогруппы R, а также их производных гаплогрупп R1, R1a и R1b, тысячелетия спустя ушли в Европу, в которую пришли в разные времена – гаплогруппа R1a прибыла в Европу через Балканы примерно 8-10 тысяч лет назад, гаплогруппа R1b прибыла в Европу 4800 лет назад через Пиренеи, и примерно 4500 лет назад через Апеннины, Балканы, из причерноморских степей, и севернее, видимо, через территорию современных Польши-Германии. Никакого «потока генов» в Америку от них не было, и состоялось только во времена Колумба и позже.
Полагаю, этих четырех примеров достаточно, чтобы понять фундаментальное заблуждение попгенетиков. Они отказываются понимать как минимум два положения.
Первое – это работать не «линейно», проводя прямую линию между носителями геномов. Они не ощущают вклад общих предков популяций, что геномные сходства, геномные вклады идут от них к их потомкам, уже разошедшимся во времени и пространстве. Не понимая этого, попгенетики не понимают древних миграций, и как те миграции переносили геномы через всю Евразию и дальше. Попгенетики же сравнивают геномы поперек эпох, поперек тысячелетий, поперек длинных миграций. В итоге они получают полностью искаженную картину, в которой, кстати, нет и временной, хронологической компоненты. Из геномов они ее получать не умеют.
Второе положение, которое не понимают (и не хотят понимать) попгенетики – это то, что необходимо, непременно надо рассматривать гаплогруппы и гаплотипы популяций в совокупности с геномными исследованиями. Причем, если рассматриваются полные геномы (или их фрагменты, так называемые аутосомные, то есть те, которые рекомбинируются между мужчинами и женщинами), то необходимо рассматривать гаплотипы и гаплогруппы как Y-хромосомы, так и митохондриальных ДНК (мтДНК). Это – опорные колонны геномных исследований. К сожалению, отсутствие добротной научной школы у попгенетиков, о чем я неоднократно сожалел в своих очерках, их изоляционизм, низкий уровень исследований, не позволяет им это усвоить. В итоге – полный конфуз в их экзерсизах.
Об этом мы и поговорим дальше, на более глобальном примере. Название его – выявление общих предков европейцев. Остается только горестно качать головой, наблюдая, как при таком низком уровне исследований и столь фундаментальном непонимании основных принципов изучения древних популяций, их уводит все дальше в дебри абсурда.
Перед тем, как заняться общими предками европейцев, еще раз посмотрим на суть методологии попгенетиков в геномных исследованиях. Что они делают? Берут, допустим, триста французов. Как мужчин, так и женщин. При этом даже не говорится, сколько мужчин, сколько женщин, это для попгенетиков неважно. Иначе будет неполиткорректно. Заложили в компьютер эти самые сотни тысяч и миллионы снип-мутаций, по всем тремстам людям, усреднили, получили «французский геном». Так же получили «русский геном». Откуда «русский», откуда взят – это усредняющих не интересует. А взят он вот откуда, цитата из моей статьи на Переформате почти год назад:
Карта показывает точку с подписью «русские – HGDP». Сокращение означает Human Genome Diversity Panel. Это – географическое место, где по международным понятиям находится «стандартный русский геном». Данное место российские популяционные генетики в своей бесконечной мудрости поместили в Архангельскую область, с самой большой в России долей финно-угорского населения. Вот доли гаплогруппы N1 в тех местах, и уже не южно-балтийской, а финно-угорской ветви:
• Мезень – 53%
• Красноборск – 40%
• Пинега – 40%
И это при том, что в среднем по европейской части России доля гаплогруппы N1 составляет 14% (и то за счет перевеса N1 севернее Пскова и Новгорода), а в центральном и южном регионах России – менее 10%. Короче, столь бестолковым выбором места для «стандартного генома русских» для международной общественности, попгенетики одним росчерком пера записали всех русских в финно-угры. И это уже не изменить, это стало официальной информацией от России.
Я не к тому, что быть финно-угром – плохо, вовсе нет. Я к тому, что эта непрофессиональность российских попгенетиков уже стала наносить открытый вред научным представлениям, которые должны быть честными и обоснованными. Она, эта непрофессиональность, исказила «генетический профиль России» во всех текущих и будущих генетических исследованиях в России и за рубежом.
Короче, русские, благодаря полному отсутствии квалификации матери и сына Балановских, ведущих попгенетиков России, которые эти данные подавали, в геномной базе данных уже приравнены к угро-финнам. Поэтому везде, где в геномных работах звучит «русские», надо читать – «Не русские, а угро-финны. По русским данных нет». Я еще вернусь к этому ближе к концу этой критической статьи.
Я прошу читателя понять, почему я принимаю иронический тон, говоря об изысканиях популяционных генетиков. Но по-другому нельзя, потому что анализ их изысканий может проводиться только в критическом ключе. Я понимаю, что это у них наука такая. Берется одно множество аутосомных мутаций (это где мужские и женские перемешаны, составляя чудовищную кашу), называется «северным евразийским», затем берется другое множество, называется «европейским», из одного множества вычитается другое, получается из одной каши вычесть другую кашу, потом из этого всего вычитаются мутации шимпанзе. Так сказать, фильтруется. Проблема в том, что шимпанзе берется один, а уже сейчас известны их разные геномы; то есть геномы в принципе близкие, но с вариациями. Естественно, при вычитании вариации уже не учитываются, вычитается какой-то один из геномов шимпанзе. Все понимают, что из разницы каш вычитается нечто, что вариабельно. Какой-то смысл в этом может быть, только смысл очень плавающий. Этому можно только посочувствовать, ну, наука у попгенетиков такая, пока только в начальной стадии развития. Многого от нее ожидать не приходится.
Но проблема вовсе не в этом. А в том, что после всех вычитаний каш из каш и еще вычитают маленькую тележку, вдруг объявляется, что вся Европа образовалась из трех популяций. Которые тоже составили и обработали подобными методами, то есть это еще три каши. Но так посчитал компьютер. И это объявляется результатом исследования. О том, что это все каша, не говорится. А порой вообще, как в случае с русскими, вместо одной популяции подсовывается другая. И что самое занятное, эту кашу редуцируют до точных цифр, процентов, долей. Вот так и оказалось, что в этнических русских 11.7% американских индейцев, о чем я уже излагал. А на самом деле никакие это не американские индейцы, это снип-мутации от общих предков, например, гаплогруппы Р, и цифра процентов на самом деле может быть любая. Но так посчитал компьютер. На него можно свалить все, что угодно.
Итак, последние несколько дней я разбирался с тем, что популяционные генетики продолжают вытворять с геномными исследованиями. Только что вышла статья про три древних популяции, которые якобы породили всех современных европейцев («Ancient human genomes suggest three ancestral populations for present-day Europeans»), под которой подписалось 98 авторов. Такое количество – уже феномен, и можно только догадываться, как это могло получиться. Создаётся впечатление, что пешкам страстно надо куда-то прицепиться и иметь еще одну публикацию. Начальникам же важно показать, сколько у них пешек-солдат. Всё равно, что мечение территории, как у собак или волков. Отобраны «наши», а кто не отобран – те не «с нами». Поэтому я сделал критический разбор этой статьи, у которой 98 авторов, а дитя, как говорится, без глазу…
Мой очерк можно было бы назвать «Берегитесь популяционных генетиков в исторических исследованиях», как называлась глава 16 в книге «Происхождение славян» (Издательство Алгоритм, Москва, 2013, 511 стр.), а до этого также называлась серия статей в Вестнике ДНК-генеалогии.
Итак, очередная статья по анализу генома попгенетиками. Это уже означает как данность – будет очередной цирк. Но эта статья – знаковая. В ней почти сто авторов. Просто так в статьях по сто авторов не бывает. Это значит, статья заявлена как эпохальная. Еще бы – найдены предки жителей современной Европы. Три предковые популяции. И об этом ста авторами объявлено на весь мир.
А откуда сто авторов-то? Так тоже не бывает. Так раньше никогда не было. Вы видели у Эйнштейна, Менделеева, Паскаля по сто авторов в статье? Да и вообще – как сто авторов могут писать статью?
А они ее и не писали. Как поясняет сноска к статье, ее писали три автора, а по сути – только один, Иосиф Лазаридис, сотрудник Гарварда и МТИ. Еще двое, как начальники, статью просмотрели, возможно, сделали замечания – это David Reich из Гарварда и Johannes Krause из Германии. Некоторые писали отдельные части Приложения к статье. Остальные соавторы, типа все тех же Балановских, присылали образцы ДНК. Это полезно, сомнений нет, только за такую техническую работу раньше в соавторы не вставляли, а выносили благодарность в сноске. Это, конечно, мелочь, но показывает стиль современной «науки».
Да, надо сказать, что сама основная статья, с основными выводами, занимает 8 страниц текста, плюс имеются три приложения, из которых только одно занимает больше ста страниц. Так что же в основной статье приведено? Разберем по пунктам.
Для начала – трех исходных популяций 8000-7500 лет назад, о чем в статье идет речь, для древней Европы явно мало. Одних мужских гаплогрупп было больше – это были I1, I2, J2, G2a, E1b-V13, R1a – и это как минимум. Были наверняка и F, и K. Из женских гаплогрупп – были U, U2, U4, U5, U8, N1a, T2, K, J, HV, V, W, X. Каждая из них – древняя, уходящая корнями на десятки тысяч лет назад. Из статьи совершенно неясно, что авторы понимают под «популяцией», но если это мужские гаплогруппы, то их минимум уже шесть, а если женские – то минимум двенадцать. И каждая корнями уходит опять на десятки тысяч лет. А если популяция – это сочетание мужской и женской гаплогруппы, так их уже десятки. Как там могли оказаться всего три – это вообще какая-то ерунда. Но так получилось. Так усреднил компьютер.
Но у попгенетиков своя логика. Три популяции, оказывается, это те три, которые они рассматривали. Рассмотрели бы пять – стало бы пять общих предков, правда, опять было бы непонятно, как они «общих предков» рассматривали. Дай им современную Москву, так у них было бы четыре предка города – чеченцы, азербайджанцы, украинцы и русские. Возможно, добавили бы таджиков и китайцев. Вот так работает популяционная генетика в исполнении ее сегодняшних творцов. Так какие исходные «популяции» они рассматривали? Действительно, поначалу всего три:
(1) женщина из раскопок в Штутгарте (современная Германия), археологическая культура линейно-ленточной керамики (LBK) датировка ∼7500 лет назад, мтДНК гаплогруппа Т2;
(2) мужчина из раскопок в Лошбуре (современный Люксембург), датировка ∼8000 лет назад, мужская гаплогруппа I2a, мтДНК гаплогруппа U5;
(3) семь мужчин из раскопок в Мотале (Швеция), датировка ∼8000 лет назад, мтДНК гаплогруппа U2, хотя в статье это было сказано невнятно («Loschbour and all Motala individuals belonged to haplogroups U5 and U2» – то ли Лошбур имел U5 и Мотала U2, то ли они все имели U5 и U2 вразнобой; подобная невнятность для статьи очень характерна); из этих семи человек пятеро были мужчинами, из них у четверых была гаплогруппа I, у пятого гаплогруппа дала противоречивые результаты; какая гаплогруппа была у двух оставшихся женщин – опять непонятно, то ли U2, то ли U5.
По этим данным, гаплогруппа I (скорее I2a, см. ниже) явно доминировала в Европе 7500-8000 лет назад, на примерах Люксембурга и Швеции. Заодно узнали, что ни женщина из Штутгарта, ни мужчина из Люксембурга не могли усваивать молоко, у обоих были темные волосы, у женщины были коричневые глаза, а у мужчины из Люксембурга, с гаплогруппой I, была 50%-я вероятность иметь голубые глаза. В общем, ни для чего последующего эта информация не была нужна, ну да ладно. До кучи сгодится.
Еще занятная информация – женщина из Штутгарта возрастом 7500 лет назад «кластеризуется» с тирольским «ледовым человеком Отци» (гаплогруппа G2а) 5300 лет назад (сейчас это пересчитано на 4550 лет назад – ААК) и с «фермером» из Южной Швеции 5000 лет назад (гаплогруппа не приведена). Люксембуржец гаплогруппы I «кластеризуется» с «охотником-собирателем» из Испании 7000 лет назад (авторы не сообщают, что у того была гаплогруппа E1b-V13), и ископаемые носители гаплогруппы I из Швеции «кластеризуются» с неким древним «шведом» возрастом 5000 лет (гаплогруппа не приведена). Эти кластеры и были названы «метапопуляциями» под названиями, соответственно, «ранние европейские фермеры», «западноевропейские охотники-собиратели» и «скандинавские охотники-собиратели».
Геномы всех трех древних людей или групп людей, определенные в статье (женщина, «люксембуржец» и «шведы»), плюс несколько геномов древних ископаемых людей, определенные ранее (среди них два сибиряка, 24 и 17 тыс. лет назад, которых окрестили «древними северными евразийцами», а также два генома с Пиренейского полуострова, геном «ледового человека» Отци, и еще один древний шведский «фермер») сравнивались с 2196 геномами 185 современных популяций, у которых были определены около 600 тысяч снипов по всем хромосомам.
Авторы пришли к выводу, что вся современная Европа порождена тремя предковыми группами. Одна – «древние северные евразийцы», которые близки сибирякам верхнего палеолита, по соображениям авторов. Что за «сибиряки верхнего палеолита» – из основной статьи непонятно, то ли R (но в древней Европе таких не было), то ли Q (но в древней Европе их тоже не было), то ли, скорее всего, женская гаплогруппа U. Но как древняя Европа могла быть «порождена» женской гаплогруппой, какая бы они ни была, и тем более, когда ей десятки тысяч лет – остается совершенно непонятным. А как насчет мужчин? Или они не считаются? Странно, что никто из 95 соавторов статьи такого вопроса не задал тем, кто статью писал.
Из приложения к статье (но не из основной статьи) выяснилось, что в состав «древних северных евразийцев» входит древний носитель гаплогруппы R, тот самый мальчик из Мальты. Но авторы не знали, судя по расчетам и обсуждению, что этот древний носитель гаплогруппы R тогда, 24 тысячи лет назад (когда он жил) в Европе не был, как не были и его предки гаплогруппы R. Поэтому «северные евразийцы» в его представительстве не могли войти в состав тех, кто «породил современную Европу». Это были его далекие потомки, пришедшие в Европу почти 20 тысяч лет спустя. Тогда они «северными евразийцами» давно не были.
Наверное, авторам не стоило бы запутывать картину с «древними северными евразийцами». Что они нашли – это, по сути, следующее: в геномном формировании популяций древней Европы (7500-8000 лет назад) приняли участие, в частности, древние гаплогруппы U (митохондриальная) и I2 (Y-хромосомная), около 5000 лет назад в Европу прибыли носители гаплогруппы R c субкладами, и они в совокупности сформировали большинство современного населения Европы. Предки гаплогруппы R сделали в далекой древности огромную миграционную петлю от того места, где они возникли как гаплогруппа ВТ примерно 60 тысяч лет назад, прошли через Южную Сибирь примерно 40-20 тысяч лет назад, и частью прошли в Европу в интервале 10-4 тысяч лет назад. Сибирь – это важный транзитный пункт в их древних миграциях.
Но поскольку авторы статьи не рассматривали ни древние миграции, ни (по сути, «функционально») гаплогруппы, ни тем более гаплотипы, и потому не рассматривали хронологию древних миграций, то их трактовки в большинстве случаев были обречены на запутанность и неверность. Проводилось формальное, «линейное» сопоставление массовых компьютерных расчетов поперек миграций, времен, эпох.
Вернемся к «популяциям», предложенным авторами статьи для рассмотрения. Вторая «популяция» – западноевропейские «охотники-собиратели», родня тому, кто из Люксембурга. Для меня всегда загадка, почему попгенетики всегда обозначают, кто там были «охотники-собиратели», а кто «фермеры». Откуда они это знают? А ответ прост: ранее 8 тысяч лет назад – это «охотники-собиратели», а позже – «фермеры». То есть датировка уже об этом полностью говорит, во всяком случае, попгенетики так считают. Зачем тогда писать эту тавтологию – «охотники-собиратели» или «фермеры», когда на самом деле они просто смотрят на датировку? Загадка мироздания. А что, если это был древний стеклодув? Отнести его к «охотникам-собирателям» или «фермерам»? А если военный был, солдат? Он кто – охотник-собиратель или фермер? Вот такие бездумные штампы у попгенетиков просто забавляют. В любом случае, по данным авторов статьи, эти из района Люксембурга породили европейцев, но не ближневосточных людей. Уже хорошо. Но в любом случае в понятиях гаплогрупп, «западноевропейские охотники-собиратели» – это опять носители гаплогруппы U (митохондриальная) и гаплогруппы I (Y-хромосомная), только без наложения более поздней гаплогруппы R.
Неудивительно, что группа «шведов» (гаплогруппы I+U) идут на диаграммах авторов (в Приложении) вместе с «люксембуржцем» (гаплогруппы I+U). Они вместе и есть «западноевропейские охотники-собиратели». Гаплогруппа U (мтДНК) с тех пор в Европе в значительной степени пропала (за редким исключением ее сейчас там единицы процентов), заместившись в основном гаплогруппой Н (мтДНК), которой в Европе сейчас около 50%. Для иллюстрации, в Швеции сейчас гаплогруппы U 2.8%, U2 0.8%, U3 0.6%, U4 3.0%, U5 12.1%. Гаплогруппы Н в Швеции – 48%. Во Франции те же гаплогруппы составляют сейчас, соответственно, 1.4%, 1.6%, 1.0%, 2.5%, 8.2%. Гаплогруппы Н во Франции – 47%. А мужской гаплогруппы I в Швеции сейчас 42%, во Франции – 15%. Но ничего этого читатель в рассматриваемой статье не найдет, хотя это и имеет самое прямое отношение к тому, кто были предковыми популяциями современной Европы.
Третья «популяция» – «ранние европейские фермеры» (можно подумать, попгенетики знают, фермеры они были или нет; бездумные штампы продолжаются), в основу которых была взята ископаемая женщина из Штутгарта. Она и есть «фермер». Она, по мнению (или данным?) авторов имела ближневосточное происхождение, но была также близка к геному ископаемого человека из Люксембурга, который к Ближнему Востоку отношения уже не имел. Как мы видим, интрига (или абсурд) уже закручиваются. И это вынесено в Абстракт статьи (!). Абстракт заканчивается тем, что 44% генома «раннего европейского фермера», той, которая из Штутгарта, происходит из «базальной Евразийской линии», которая отщепилась «перед разделением неафриканцев». Кто что понял? Я – нет. Что такое «базальная Евразийская линия»? Зачем такие уклончивые пассажи? Если бы это была мужская гаплогруппа, например, P, то она по геному легко определяется. Это – любой из снипов 92R7_1, 92R7_2, L138, L268, L405, L471/PF5989, L536/PF5860, L721/PF6020, L741, L768/PF5976, L779/ PF5907/YSC0000251, L781/PF5875/, M45/PF5962, M74/N12, P27.1_1/P207, P27.1_2, P69, P226/PF5879, P228/PF5927, P230/PF5925, P235/PF5946, P237/PF5873, P239, P240/PF5897, P243/PF5874, P244, P281/PF5941, P282/PF5932, P283/PF5966, P284, P295/PF5866/S8, Page83, V231.
Например, снип 92R7_1 – это мутация цитозина в аденин в положении 9,105,720 в Y-хромосоме ДНК. И тогда прямо сообщали бы – гаплогруппа Р, а не это вязкое «базальная Евразийская линия», что ровным счетом ни о чем не говорит. К тому же это у женщины из Штутгарта, и что это у нее обозначает? Как это – «перед разделением неафриканцев». Что это вообще такое? Они же начали разделяться примерно 50-60 тысяч лет назад, а здесь – 7500-8000 лет назад. Из статьи – совершенно непонятно. Какой-то птичий язык. Опять – где были остальные 95 соавторов, которые бы сказали – зачем ерунду писать? Либо мы знаем, либо мы не знаем. В каждом случае надо так и писать.
Хорошо, переходим к тому, что, как оказалось, у всей Европы есть только три предковых популяции, причем всего 7500-8000 лет назад, причем почти все обнаруженные мужские линии – гаплогруппы I, причем как нарочно именно те три, которые раскопали и взяли за референсные ДНК-линии. И что такое вообще – предковые популяции, когда речь идет о популяциях мужчин и женщин, которых 7500-8000 лет назад должно было быть много, в разных комбинациях мужских и женских гаплогрупп. И куда делись мужские гаплогруппы G2a, R1a, E1b, J2? И много других женских, найденных в раскопках того же времени? Ясно, что здесь должно быть какое-то огромное приближение («огромное приближение» – это здесь эвфемизм понятию «каша»), и какой в этом смысл?
Давайте разбираться, хотя разобраться в каше невозможно. Статья написана очень вязким языком, фразы уклончивы – ясно, что ничего определенного там нет. Но – почти сто авторов. Какая-то фантасмагория.
Вот – пример: «…the ancestors of Stuttgart were not unmixed migrants from the Near East». Что авторы хотели сказать, используя неуклюжее not unmixed? Видимо, что предки женщины из Штутгарта были смесью Ближнего Востока с чем-то еще, а с чем – не пояснено. Это что – информация? Чтобы не подумали, что я вырываю это из контекста, расширим цитату и приведем пояснения. Авторы использовали PCA (Principal Component Analysis), и написали, что это «мощный» метод, но «интерпретация получаемых результатов в отношении истории сложна». Это ясно, все ДНК-данные непросто интерпретировать в отношении исторической панорамы. Поэтому авторы заранее поставили вопрос – можно ли определить, что современные европейцы образовались от трех древних популяций – Loschbour (это – современный Люксембург), Штутгарт (современная Германия), и некая таинственная популяция MA1 (про таинственность – см. далее). Что такое МА1 – авторы просто забыли указать в основной статье. Немудрено, когда 98 авторов. На самом деле это тот самый древний мальчик из Мальты. Древних шведов в этой тройке уже забыли. Действительно, цитируем: «if present-day Europeans were formed by admixture of populations related to Loschbour, Stuttgart and MA1».
То есть предки европейцев были уже зафиксированы с самого начала, осталось только найти обоснования, что при работе с кашами – вовсе не сложно. Что такое МА1 – авторы в тексте, повторяю, не пояснили, и опять пришлось продираться через ребусы. На странице 8 статьи авторы написали, что «we modeled Europeans as a three-way mixture of ANE (of which MA1 is a member), WHG (Loschbour), and EEF (Stuttgart)». То есть эта загадочная МА1 относится к ANE, что есть Ancient North Eurasian. WHG, что есть ископаемый из Люксембурга с гаплогруппой I – это West European Hunter-Gatherer, а женщина из Штутгарта – это Early European Farmer. Далее, говоря об МА1, авторы написали, что «Native Americans are genetically closer to MA1 than to European hunter-gatherers», то есть что американские индейцы более близки генетически к этой МА1, чем к европейским охотникам-собирателям (дались эти «охотники-собиратели» авторам, когда это на самом деле просто древние европейцы ранее 8000 лет назад), и ясно, что американские индейцы к ним не близки, как не близка гаплогруппа Q к гаплогруппе I. А вот как эта МА1, которая относится к древним северным евразийцам и близка к американским индейцам, вообще появилась у авторов, они в основной статье не поясняют. Это потому что у нее может быть мтДНК гаплогруппа U? Так у американских индейцев ее нет, у них в основном A, B, C, D, и X, да и Х всего около 3%.
Потом авторы раскрывают, что это было просто исходное предположение. Авторы так и пишут – «we developed an independent method for estimating mixture that only assumes that MA1 is a representative of ANE, Loschbour of WHG, and Stuttgart of EEF». То есть авторы assume, закладывают исходное допущение, что МА1 представляет древних северных евразийцев, «Люксембург» с гаплогруппой I – древних западноевропейцев (забудем про «охотников-собирателей» как примитив), и древняя женщина – менее древних (то есть более современных) европейцев же (забудем про «фермеров» по той же причине). Положить этих трех в основу всех европейцев с самого начала – это было исходное допущение. Шведов опять здесь авторы в расчет не взяли как отдельную популяцию, но по всему тексту идет, что они связаны с «древними северными евразийцами», которые близки американским индейцам. Почему так – авторы не поясняют. Кто такие МА1 – в статье опять не поясняется, а основная статья почти завершилась.
Тем временем, оказалось, что «ANE ancestry was already present in at least some Europeans (Scandinavian hunter-gatherers) by ∼8,000 years ago», то есть предки ANE (Ancient North Eurasian, близкие к американским индейцам), уже оказываются предками скандинавов (опять «охотников-собирателей») 8 тысяч лет назад, у которых (скандинавов) обнаружена мужская гаплогруппа I. Это опять потому, что у них могла оказаться женская гаплогруппа U? Ну, знаете, при таком числе вариаций можно все на свете объяснить.
А откуда появилось, что древние северные евразийцы оказались предками скандинавов 8 тысяч лет назад? А вот почему – «ANE ancestry was already present in at least some Europeans (Scandinavian hunter-gatherers) by ∼8,000 years ago, since MA1 shares more alleles with Motala12 than Loschbour: f4 (Motala12; Loschbour; MA1, Mbuti) = 0.003 (Z=5.2 standard errors from zero)». Всем понятно? Это потому, что таинственная МА1 имеет «больше общих аллелей со скандинавами (Motala12), чем с люксембуржцем», хотя оба последних – гаплогруппа I, а кто такая МА1 – мы так пока и не знаем, кроме того, что это часть северных евразийцев, близких к американским индейцам (в Приложении делаются намеки, что это – мальчик из Мальты, хотя слово «Мальта» или «Байкал» не употребляют). Ребус продолжается. Неплохо, не так ли? И все это получено с помощью так называемой f-статистики. Как поясняют авторы, f3 – это сопоставление образца с двумя популяциями в отношении частот определенных нуклеотидов, а именно (X-Ref1) и (X-Ref2). Авторы сопоставляли все современные европейские геномы с каждым из четырех геномов, а именно «люксембуржца», женщины из Штутгарта, шведов (Мотала12) и этого таинственного МА1. Последний в основной статье так и не определен (у 98 авторов дитя без глазу), только в подписи к рис. 1 мелькнуло следующее «MA1 falls outside the variation of modern day West Eurasians in the direction of southern-northern differentiation along dimension 2 and between the European and Near Eastern clines along dimension 1». В общем, пусть кто как хочет, так и разбирается.
Говоря о вязком языке статьи, вот – типичный пример, в котором и f3 статистика затронута. Цитирую: « While SHG may have contributed ANE ancestry to modern Europeans, it cannot have been the only population that did so, as no European population has its lower f3-statistic with it in Table 1, and few populations fit a model of EEF-SHG admixture».
Всем всё понятно? Расшифровываю: «В то время как шведские «охотники-собиратели» могли донести вклад древних северо-евразийских предков в (геном) современных европейцев, они не могли быть единственными, кто это сделали, поскольку нет такой европейской популяции, которая имела бы с ними более низкую f3-статистику в таблице 1, и почти никакая (или просто никакая) популяция не соответствует примесной модели ранних европейских фермеров (это – женщина из Штутгарта с Отци и «шведом» – ААК) со скандинавскими охотниками-собирателями».
Яснее стало? И вот такой птичий язык – по всей статье. Проверить что-либо совершенно невозможно, как, впрочем, и понять. Не помогает и табл. 1, в которой нет ни одной цифры. Это просто некое перечисление стран, для которых сопоставление геномов дало «наиболее негативное f3» с двумя древними геномами – «люксембуржца» и женщины из Штутгарта. Древних «шведов» с гаплогруппой I в таблице 1, которая на самом деле в основной статье – единственная таблица, уже нет. Посмотрим в таблице на «русских» (что на самом деле финно-угры, см. выше). Впечатляет. «Наиболее негативное f3», что, если следовать авторам статьи, «if X is admixed between populations related to Ref1 and Ref2, the statistic can be negative and provides evidence of admixture in population X5», то есть те страны, которые примесные с «люксембуржцем» и странами Ближнего Востока, которые состоят из Абхазии, Армении, Грузии, и иракских евреев, наблюдается для: Эстонии, Финляндии, Исландии, Литвы, Мордовии, России, Белоруссии, Чехии, Норвегии, Украины, басков, Англии, южной Франции, северной Испании, Испании и Шотландии.
Опять – всем все понятно? То есть эти страны admixed между носителем древней Y-хромосомной гаплогруппы I и мтДНК U5 с территории современного Люксембурга, и кавказцами наряду с иракскими евреями. Действительно впечатляет, не так ли? Правда, авторы написали – «we observe that Finns, Mordovians, Russians, Chuvash, and Saami from northeastern Europe do not fit our model». То есть – «мы получили, что финны, мордва, русские, чуваши и саами не вписываются в нашу модель». Замечаете, в какой группе находятся русские? Естественно, в той самой группе, в основном, финно-угров, куда и записали русских попгенетики Балановские, подав неверные данные по геномам в международную базу данных.
Вот и карта из статьи, которая это показывает:
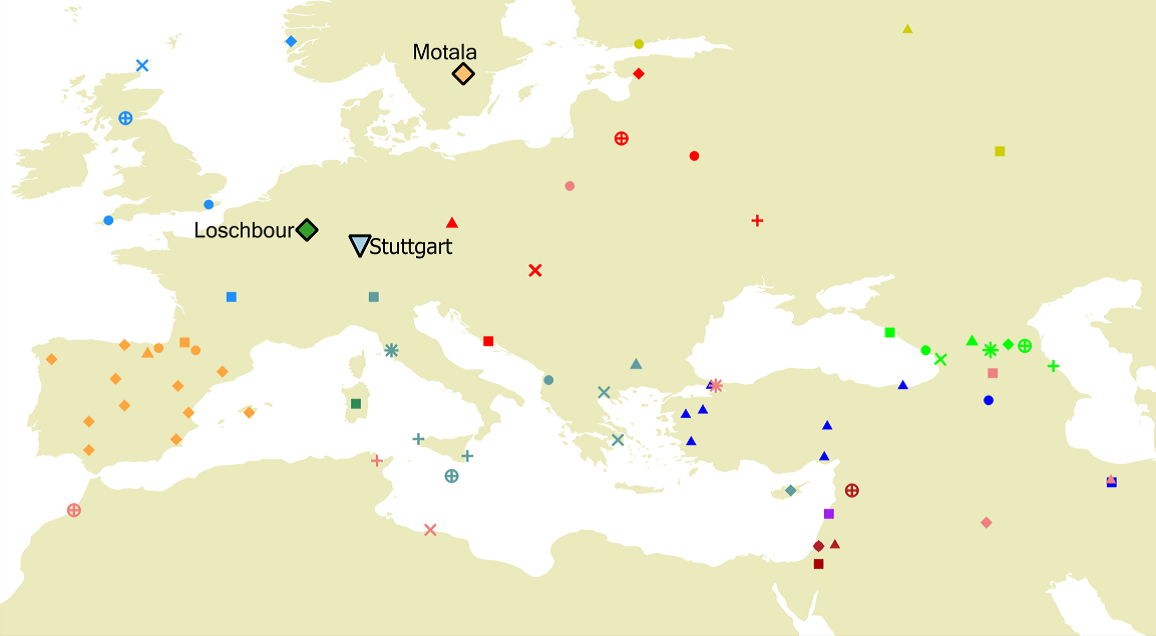
Хотите знать, где на этой карте располагаются «русские»? Вот где – маленький зеленый треугольничек в верхнем правом углу карты, северо-восточнее Балтийского моря. Севернее мордвы. Намного восточнее Финляндии. Вот именно из этого места российские попгенетики сдали образцы для определения генома, выдав это за «русский геном». Теперь в геномной базе данных Россия – там. Видимо, навсегда. Кто теперь это будет менять? Все и так знают, что «русские – это угро-финны», и попгенетики Балановские это подтвердили. Какие еще могут быть вопросы? Балановские же ведь не признаются, что совершили, по сути, научное преступление.
Что там еще авторы насчитали такими методами? А вот что:
(1) Жители Океании, Восточной Азии, американские индейцы и островитяне Бенгальского залива между Индией и Бирмой «генетически более близки» к древнему люксембуржцу, древним «шведам» из Моталы и пока все еще неясному МА1, чем к женщине из Штутгарта.
(2) Они же (исключая американских индейцев) генетически близки, причем «в равной степени», все к тем же люксембуржцу, «шведам» и МА1, но американские индейцы более близки к МА1, чем к «европейским охотникам-собирателям».
(3) Все трое «охотников-собирателей» плюс женщина из Штутгарта (она, напомним, «фермер») генетически более близки к американским индейцам, чем к перечисленным выше жителям Океании, Восточной Азии и тем самым островитянам.
Кто что понял в этом «генетически более близки» и какой в этом смысл в отношении происхождения европейцев, о чем намеревалась рассказать нам статья? И вот такой вязкий стиль – по всей статье.
Статья заканчивается так: «This study raises two questions that are important to address in future research. A first is where the EEF picked up their WHG ancestry. Southeastern Europe is a candidate as it lies along the geographic path from Anatolia into central Europe, and hence it should be a priority to study ancient samples from this region. A second question is when and where ANE ancestors admixed with the ancestors of most present-day Europeans. Based on discontinuity in mtDNA haplogroup frequencies in Central Europe, this may have occurred during the Late Neolithic or early Bronze Age ∼5,500-4,000 years ago. A central aim for future work should be to collect transects of ancient Europeans through time and space to illuminate the history of these transformations».
Краткий перевод: «Наше исследование поднимает два вопроса. Первый – где «ранние европейские фермеры» (то есть это женщина из Штутгарта вкупе с Отци гаплогруппы G2a и древним «шведом» гаплогруппы I – AAK) заполучили свою родословную западноевропейских охотников-собирателей? Кандидат на это – юго-восточная Европа, и оттуда надо изучать образцы (ДНК). Второй вопрос – когда и где древние «северные евразийцы» смешались с предками большинства современных европейцев? Основываясь на исчезновении мтДНК в центральной Европе (в отношении частот их гаплогрупп), это могло произойти 5500-4000 лет назад. Центральная задача будущих исследований состоит в сборе данных по древним европейцам в пространстве и во времени».
Читать это несколько странно. Не случайно, видимо, авторы вообще не касались мужских гаплогрупп ископаемых скелетных останков, как, впрочем, и женских, только приведя их в начале статьи. А эти гаплогруппы, в первую очередь, и указывают на родословные, на общих предков популяций. Авторы же их вообще не обсуждали. Такое ощущение, что тот, кто статью писал, в гаплогруппах вообще не разбирался, а остальные просто статью подписали. Впрочем, сейчас в редакциях и подписи соавторов не требуются. А у изучаемых древних людей в основном гаплогруппы I, что, конечно, вовсе не исчерпывает гаплогруппы Европы 7-8 тысяч лет назад. Гаплогруппа I, вполне возможно, пришла с Русской равнины, или, как вариант, возникла в Европе. Если пришла с Русской равнины, то, вполне возможно, в паре с гаплогруппой U. Но этим, конечно, Европа 7-8 тысяч лет назад вовсе не исчерпывается. Искать предков по аутосомам, да еще в столь примитивном варианте, как это делали авторы – путь практически безнадежный. Игнорировать гаплогруппы – тоже путь безнадежный. Анализировать «геномы этносов» можно только в самом первом, если не в нулевом приближении. Задачу это не решит.
Ответ на вопрос «когда и где древние северные евразийцы смешались с предками большинства современных европейцев» (см. выше) уже давно получен. Большинство современных европейцев-мужчин относятся к гаплогруппе R1b, так что данная статья их и не касалась, поскольку они пришли в Европу относительно недавно, всего около 5000 лет назад. Там и смешались с предками европейцев «Старой Европы», причем в основном с женщинами, преимущественно гаплогруппы Н. В статье они тоже не затрагивались. Так что сам «ответ» про три популяции как предковые современных европейцев просто неверен.
И это – вклад в науку 98 авторов статьи. Ну, и какая была мотивация этих авторов войти в состав этой статьи, кто пояснит? Я могу пояснить, но оставлю это нелицеприятное мнение при себе. Впрочем, выше отчасти написал.
Настала, наверное, пора перейти к приложениям к статье с их объемом более чем в сотню страниц, но этого делать не стоит. И вот почему. Ценности в них нет никакой. В основу статьи, да и всего подхода вообще, положено фундаментальное заблуждение. Подход основан на формальном «линейном» сопоставлении десятков и сотен тысяч снип-мутаций, которые на самом деле образуют не линейную систему, а «трехмерную», условно говоря, где важной является хронологическая компонента. Хронологию появления и миграций снип-мутаций авторы вообще не учитывают. То есть они цитируют времена древних ископаемых образцов, но это не та хронология. А в реальности – там и мужские, и женские снип-мутации, которые наслаивались друг на друга и перемешивались десятки тысяч лет. Миграции этих мутаций происходили по разным направлениям, порой расходясь из одного региона, другие через этот регион проходили в разные исторические эпохи, вплоть до недавнего времени. Поэтому авторы, рассматривая массивы мутаций из разных регионов у современных людей, зачерпывают пласты снип-мутаций за десятки тысяч лет. Никаких определенных «общих предков» там нет, там напластования суперпозиций снип-мутаций. Более того, авторы полностью игнорируют гаплогруппы и гаплотипы рассматриваемых популяций, и ископаемых людей. А эти гаплотипы-гаплогруппы, как Y-хромосомные, так и митохондриальные, являются опорными колоннами, которые приводят в определенную систему напластования снип-мутаций. Авторы, повторяю, это опорные колонны полностью игнорируют, и в анализе не рассматривают.
Чтобы было понятнее, о чем речь, рассмотрим ту самую неуловимую (по статье) популяцию (или представителей) МА1. Авторы почему-то окружили ее завесой секретности. Наверное, ясно почему – она ломает большинство их построений, и показывает практическую абсурдность подхода. Напоминаю, что в основном тексте статьи МА1 не была расшифрована, и намеки о ее происхождении, опять в виде ребуса, появляются только в конце одного из Приложений.
Как уже мы догадались и сообщили выше, МА1 – это ребенок из поселения Мальта, рядом с Байкалом, с датировкой 24 тысячи лет назад. У него найдена Y-хромосомная гаплогруппа R и мтДНК гаплогруппа U. Но 98 авторов и про это не пишут, гаплогруппы не раскрывают, слова «Мальта» не упоминают. Вот как они обозначают в Приложении эту важную находку, и важную именно потому, что у нее есть привязка к гаплогруппе R: «the recently published Paleolithic MA1 sample from Siberia». И еще: «the time of MA1 (∼24,000 years ago)». И, что то же самое, «MA1 is an Upper Paleolithic Siberian».
Всё. Больше ничего конкретного о ней не открывают. Но общих разговоров о ней у авторов много, и все невпопад. Сейчас поясню, почему. Это – хороший пример иллюстрации сюрреализма 98 авторов статьи. Выше я уже это пояснял, но сейчас расскажу другими словами, поскольку это важная часть абсурда, разворачиваемого авторами статьи.
Итак, гаплогруппа R была найдена в Южной Сибири. Она образовалась из родительской гаплогруппы Р, и там же (видимо) образовался ее «брат», гаплогруппа Q. Поэтому их геномы должны быть очень сходны. Гаплогруппа Q в значительной (или в заметной) степени ушла в Америку и стала американскими индейцами. Гаплогруппа R продолжала производить новые нисходящие гаплогруппы – R1, R1a, R1b, которые в значительной степени ушли в Европу много тысячелетий назад (R1a пришла в Европу 8-10 тысяч лет назад, R1b – около 5 тысяч лет назад), R замечена, в частности, на Кавказе, да и вообще должна быть рассеяна по всему миграционному пути из Южной Сибири, как и гаплогруппы R1a и R1b, которые до сих пор встречаются и в Сибири, и среди уйгуров, и среди турок, и вообще по всему пути вплоть до Европы, и, разумеется, в Европе, где R1a занимает половину Восточной Европы, а R1b – более половины Западной Европы. Иначе говоря, гаплогруппы R и Q разошлись в диаметрально противоположные стороны, но имели очень близкие геномы.
Теперь вопрос – будет ли геном древней гаплогруппы R перекликаться с современными геномами Европы? Разумеется. Но не с ископаемыми гаплогруппами 8 тысяч лет назад, потому что гаплогруппа R пришла в Европу только около 5 тысяч лет назад. Будет ли геном древней гаплогруппы R перекликаться с геномами американских индейцев с гаплогруппой Q (основной среди америндов)? Разумеется, это гаплогруппы-братья, от одного отца – гаплогруппы Р. Будет ли геном древней гаплогруппы R перекликаться с кавказскими популяциями? Разумеется, там, на Кавказе, много субкладов гаплогруппы R. С ближневосточными? Разумеется, там много субкладов гаплогруппы R. Как видим, все упомянутые геномы будут пересекаться, и компьютер, естественно, найдет много общего в них, и покажет, что они все «родственны». Можно и формальные проценты посчитать, даже по одним мужским гаплогруппам и геномам.
А женские? То же самое. Гаплогруппа мтДНК U тысячелетиями распространена по всей Европе, от Атлантического океана до Тихого. Это – основная палеолитическая митохондриальная гаплогруппа. Это – ископаемая гаплогруппа древней Европы (с датировками 7000-8000 лет назад), она же (гаплогруппа U2) найдена в Костенках, на Русской равнине, с датировкой около 40 тысяч лет назад. На Байкале с датировкой 24 тысячи лет назад – гаплогруппа U. В древних монгольских захоронениях – гаплогруппа U2. Теперь мы видим, что у «люксембуржца» (8000 лет назад) – гаплогруппа U5, у древних «шведов» (8000 лет назад) – U2 и U5, а геномы всех этих U, U2, U5 – перекрываются. Ну, и как удивляться, что геномы у всех перекрываются от Атлантики до Сибири? Только у женщины из Штутгарта гаплогруппа Т2, вот она, в принципе, может и не перекрываться с теми, кто описаны выше. Вот если бы те 98 авторов такую картину сначала набросали бы, не игнорируя гаплогруппы и датировки, то сразу получили бы хорошие рамки для своих интерпретаций. Но они в этом, видимо, не особо разбираются. И вот теперь посмотрим, что они «получили» в сравнении с картиной выше, для начала на примере «древнего мальчика с Мальты гаплогруппы R».
>> MA1 is an Upper Paleolithic Siberian with demonstrated genetic links to both Europe and Native Americans and thus is a powerful sample for constraining possible historical scenarios.
Это – о том, что «верхнепалеолитический» образец с Байкала (из Мальты) показывает «генетические связи» и с Европой, и с американскими индейцами. Как я уже сообщал выше, это, само собой, разумеется, поскольку тот, с Байкала – гаплогруппы R. Не случайно, видимо, авторы его гаплогруппу и не упоминают. А вот дальше – занятно: авторы пишут, что знание того, что у него связи и с Европой и с Америкой есть «мощный (инструмент) для ограничения возможных исторических сценариев». Никакого ограничения авторы, естественно, не делали, как и вообще не рассматривали серьезно никакие «исторические сценарии». Да и как они могли рассматривать? Геномы не дают ни направлений перемещений (миграций), ни датировок. А гаплогруппы-гаплотипы авторы не рассматривали. Поэтому цитата выше никакой пользы не дает. Банальность.
>> MA1 having contributed some ancestry to present-day Europeans not accounted for by West Eurasian Hunter Gatherers and Early European Farmers.
Это – о том, что «мальчик с Мальты» (точнее, гаплогруппа R и U), по мнению авторов, внес свою предковость в (геном) современных европейцев, независимо от (вклада) «охотников-собирателей» западной Евразии и «ранних европейских фермеров». Но кто такие здесь «западно-евразийские охотники-собиратели»? Опять приходится разгадывать этот вязкий ребус. Оказывается, это «a set of «West Eurasian» populations as those that cluster with Europe and the Near East in an ADMIXTURE analysis». То есть тот, кто оказался смесью европейцев с ближневосточными людьми, как насчитал компьютер по заложенной в него программе с соответствующими приближениями и допущениями. Это какие же такие ближневосточные люди, которые в своей совокупности не имеют гаплогруппы R и ее нисходящих ветвей? А так показал компьютер. Это – продолжение фундаментальной проблемы – расчет ведется формально, без понимания истории миграций, датировок, при игнорировании гаплогрупп.
Еще о «западно-евразийских охотниках-собирателях», которые на самом деле просто древние (более 8 тысяч лет назад) смеси европейцев и ближневосточных людей. А это, кстати, известно и ожидаемо, потому что гаплогруппы R1a и R1b прошли в Европу с востока через Месопотамию.
Так вот, «that European hunter-gatherers like Loschbour and Motala fall outside the variation of West Eurasians in the direction of European differentiation from the Near East. This pattern is suggestive of present-day Europeans being admixed between ancient European hunter-gatherers and ancient Near Easterners». Смысловой перевод – «европейские охотники-собиратели (дались им эти охотники-собиратели – ААК) как люксембуржец (гаплогруппа I – AAK) и древние шведы (гаплогруппа I) не входят в состав западноевразийцев. Это показывает, что современные европейцы являются смесью древних европейских охотников-собирателей и древних ближневосточных людей». Иначе говоря, сообщается, что древние европейцы не имели примесей Ближнего Востока, а современные европейцы имеют. И это совершенно понятно, знай бы 98 авторов хоть немного ДНК-генеалогию. Древние европейцы, 7000-8000 лет назад, которые имели гаплогруппу I, не имели в предках Ближний Восток. А около 5000 лет назад в Европу пришли эрбины, носители гаплогруппы R1b, со своими ближневосточными женщинами, поскольку эта миграция была долгой, между 6-7 и 5 тысяч лет назад, то есть продолжительностью одно-два тысячелетия, и принесли ближневосточные снип-мутации. То, что находят с такой вязкостью авторы, четко показано до того ДНК-генеалогией. На самом деле такое согласование – это хорошо, только с самого начала ДНК-генеалогию нужно было принять во внимание. Об этом же и следующее положение авторов:
>> The absence of a Near Eastern relatedness in all European hunter-gatherer groups but its presence in Stuttgart (Отсутствие ближневосточного вклада в европейских охотников-собирателей, но его присутствие (у женщины) из Штутгарта).
И опять оно очевидно. У тех древних европейцев, у кого гаплогруппа I («люксембуржец», «шведы»), генетической общности с Ближним Востоком нет. Нет гаплогруппы I на Ближнем Востоке. А у женщины из Штутгарта с гаплогруппой Т2 – есть. И немудрено – гаплогруппа Т2 находится на современном Ближнем Востоке и на Кавказе в количествах от 4.5% (Ливан, Ливия, Курдистан, Турция, Ирак) до 5-7% (Иордания, Палестина, Сирия) и до 9-12% (Грузия, Чечено-Ингушетия). Но надо отметить, что «оно очевидно» только для понимающих ДНК-генеалогию. На самом же деле это (курсив выше) крайне важное положение, и о его важности см. в конце моей статьи.
А как результат древних миграций гаплогруппы R1b по Азии, Кавказу, Ближнему Востоку и далее в Европу (см. выше) – у авторов и получается:
>> Evidence tying MA1 to Europe, the northern Near East and Caucasus, and south/central Asia. И это: The high affinity of the Northeast Caucasus to MA1.
Две эти цитаты гласят, что байкальский мальчик гаплогруппы R имеет геномную связь с Европой, Ближним Востоком, Кавказом и северо-восточным Кавказом, и южной/центральной Азией. Все это, самой собой, вытекает просто из сопоставления гаплогрупп и их древних миграций.
Еще одна цитата: …we do not currently know whether the signal of admixture observed in the Near East and Caucasus reflects an arrival of MA1-related ancestry from the east, or alternatively dilution of native MA1-related ancestry by an expansion of a Near Eastern population carrying Basal Eurasian admixture, associated perhaps with the expansion of Levantine/Mesopotamian early agriculturalists who seem to have influenced the Y-chromosome distribution of the region. Future studies of ancient Central Eurasians may help resolve such questions of migration timing and directionality.
Смысловой перевод: В настоящее время мы не знаем, отражает ли сигнал примеси, наблюдаемой на Ближнем Востоке и на Кавказе, прибытие предков MA1 (гаплогруппы R Южной Сибири – ААК) с востока, или, альтернативно, разбавление «предковости» нативной МА1 за счет расхождения ближневосточной популяции, переносящей базальную евразийскую примесь, которая, возможно, связана с экспансией ранних агрокультуристов Леванта/Месопотамии, которые, видимо, повлияли на распространение Y-хромосомы в этом регионе. Будущие исследования древних центральных евразийцев может помочь разрешить эти вопросы миграций во времени и по направлениям.
Всем понятно? И вот такой вязкий текст – по всей статье, что я уже не раз отмечал. И вот так – все сотня с лишним страниц Приложения, как и основной статьи. Авторы не понимают, что не предки гаплогруппы R прибыли с Востока, а, напротив, потомки. Если речь идет о предках мтДНК гаплогруппы U, то это вообще не имеет отношения к туманно задаваемому вопросу.
Так что, приходится повторить – без гаплогрупп и гаплотипов, причем как Y-хромосомной ДНК, так и митохондриальной, эту формализованную кашу никак не разрешить. И сотня авторов здесь ничем не помогут, если они не имеют понятия о том, как надлежит делать исторический анализ снип-мутаций, и что закладыванием сотен тысяч и миллионов снип-мутаций в компьютер такие задачи не решаются. А там по одному «мальчику из Мальты» гаплогруппы R в компьютер закладывалось 427 211 снип-мутаций, да по древним «шведам» из Моталы 411 453 мутации, до по «ледовому человеку Отци» 518 229 мутаций, да по ископаемому человеку с Афонтовой горы 92 486 мутаций, и так далее, но настоящего толку из этого нет и быть, по большому счету, не может. Никогда в историческом анализе формализованные расчеты не помогали. Особенно когда при них игнорируются «несущие конструкции» ДНК в виде гаплотипов и гаплогрупп, которые к тому же дают хронологию эволюции человека. К сожалению, сотня авторов статьи это, похоже, не понимает.
Неужели, скажет иной, так ничего эти 98 авторов и не нашли, при таком обилии произведенного материала? Ну почему же, нашли, конечно. В их груде материала можно найти некоторые важные находки, только авторы их проглядели и не оценили важность этих находок. На мой взгляд, наиболее важное – это то, что древние (7500-8000 лет назад) европейские носители гаплогруппы I («люксембуржец» и «древние шведы») не показали геномной связи с Ближним Востоком. Авторы пишут об этом так: «The absence of a Near Eastern relatedness in all European hunter-gatherer groups». То есть легенда, что заселение древней Европы происходило исключительно с Ближнего Востока, не подтверждается геномными данными.
Естественно, кто-то приходил и с Ближнего Востока. Это, по данным авторов, например, предки женщины из Штутгарта. Мы знаем, что наверняка оттуда приходили носители гаплогрупп J1 и J2. Пришли (видимо, на Балканы) носители гаплогруппы R1a, сами исходно не с Ближнего востока, но после пересечения Анатолии и, вполне возможно, вместе с женщинами ближневосточного происхождения. Приходили, также через Ближний Восток, и эрбины, носители гаплогруппы R1b, но это уже намного более поздние времена. Но мы сейчас говорим про гаплогруппу I, древнюю палеолитическую европейскую гаплогруппу. Если она не пришла с Ближнего Востока, о чем говорят геномные данные авторов, то для ее возникновения примерно 40 тысяч лет назад остаются наиболее вероятными только варианты Европы и Русской равнины. Соответственно, повышается вероятность, что именно там возникло современное человечество, не позднее 160 тысяч лет назад, и возникли первые европеоиды, примерно 60 тысяч лет назад. По первой дате у нас пока нет археологических свидетельств, но их отсутствие не есть невозможность возникновения Homo sapiens в Европе или на Русской равнине, хоть это пока относительно неожиданная постановка вопроса. Что касается европеоидов, то наиболее древние датировки их в Европе – 45 тысяч лет назад (Англия и Италия). Немногим менее древние ископаемые европеоиды на Русской равнине – 35-42 тысячи лет назад. В Костенках отмечены датировки и 50 тысяч лет назад, но это не скелетных остатков, а стоянок, датировка по почве.
Еще важная находка в статье (описанная в приложении отдельной секцией под двумя авторами) заключается в том, что оба древних носителя гаплогруппы I не относились к субкладу I1. «Люксембуржец» имел субклад I2а1b-M423, L178, древний «швед» из Мотала имел гаплогруппу I, не относился к I1, но на I2 его не проверяли. Еще один «швед» из Моталы оказался I2-L68. Остается под вопросом, была ли вообще гаплогруппа I1 в Европе, или она осталась, например, на Русской равнине. Данных к этому пока нет.
Интересно, что субклад I2a1b-M423 «люксембуржца» 8 тысяч лет назад – тот самый, что сейчас называют «динарским», и который наиболее часто встречается на Балканах (до 40% от мужского населения), и распространен по всей Восточной Европе, от Греции до Прибалтики, включая Украину, Белоруссию, Россию, с возрастом общего предка у всех примерно 2300 лет. Примерно 4500 лет назад он почти весь был истреблен, или, говоря более нейтрально, пропал из Европы. Как было сообщено в сетевой дискуссии по материалам обсуждаемой статьи, Ken Nordtvedt, ведущий мировой специалист по гаплогруппе I, пришел к выводу, что ДНК-линия «люксембуржца» принадлежала к ныне исчезнувшей ветви субклада I2a-M423. Так что данные ДНК-генеалогии об исчезновении линии субклада I-М423 подтверждаются независимым ДНК-анализом.
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















