
В первых двух частях этой серии статей (часть 1, часть 2) было рассказано о новых данных по ископаемым ДНК на большой территории, от Прибалтики (культура ямочно-гребенчатой керамики, нарвская культура и культура шнуровой керамики) до балканских археологических культур, с совокупными датировками от 8000-7500-7000 до 3800-3500 лет назад. Конечно, не обошлось без принципиальных сюрпризов. При этом надо понимать, что ископаемые ДНК – очень коварный объект науки. Чем менее профессиональны читатели, тем больше они придают значения единичным находкам. Они не затрудняют себя подумать, что если вырвать случайный единичный образец гаплогруппы из выборки, скажем, в современной России, то он в любом случае не будет характеризовать полигаплогруппный состав страны. Более того, он совсем не обязательно придется даже на доминирующую (в численном отношении) гаплогруппу.

Для того, чтобы понять, насколько случайно найденный образец гаплогруппы характеризует археологическую культуру, надо знать, насколько культура была однородна в этом отношении. А это не узнать без выборки хотя бы в несколько образцов ДНК. Более того, желательно иметь основательные представления о том, откуда насельники данной культуры пришли, и куда они затем (хотя бы в части) передвинулись. Что же делают откровенные дилетанты? Они выхватывают единичную находку древней ископаемой ДНК на территории, где это вообще первая находка, и публично провозглашают, что вот это и был древний народ. А поскольку провозглашают они это другим дилетантам, то вот и основа общего оглупления, которое потом транслируется в совершенно безумные комментарии в сети. С одной стороны, какая разница? Каждый сам кузнец своей глупости. Но с другой – за державу обидно.
Вот образец такой глупости. Один известный персонаж, который гуляет с форума на форум, и везде оставляет безграмотность, пишет: «Вот еще два образца по полякам, железный век и средние века, гаплогруппы G2a и I1, но ни R1a, ни R1b что-то всё так и нет». И дальше все время приговаривает, что в древней Европе гаплогруппы R1a так и не нашли. Он, видимо, не понимает несколько простых вещей – что в разных странах количество находок ископаемых ДНК совершенно разное, и что число находок ископаемой гаплогруппы R1a исчисляется десятками, гаплогруппы R1b по разным спискам или примерно столько же, или намного больше. И эти разные списки все по своему правильные. Каждый – объективная реальность.
Для иллюстрации и в качестве примеров – два списка ископаемых ДНК, один составлен в основном по данным Haak и Allentoft с сотрудниками (2015), другой – по данным группы Reich с сотрудниками (2017). В первом – 126 образцов, во втором – 416. По странам распределение образцов следующее. В первом списке:
Россия — 36 образцов ископаемых ДНК
Германия — 33
Турция — 15
Испания — 14
Венгрия — 10
Швеция — 9
Италия — 4
Дания — 3
Чехия — 1 (гаплогруппа R1b)
Польша — 1 (гаплогруппа К)
Гаплогруппы R1a среди всех 126 образцов – 25 (из них в Западной Европе 12), гаплогруппы R1b – 29 (из них в Западной Европе 11).
Во втором списке, из 416 образцов:
Западная Европа — 132 (из них 44 Германия)
Восточная Европа — 127
Балканы — 60
Иберия — 30
Анатолия — 30
Иран — 9
Италия — 6
Сибирь — 3
Кавказ — 2 (оба Грузия)
Африка — 1 (Эфиопия, 4575-4469 лет назад, гаплогруппа E1b)
Гаплогруппы R1a среди всех 416 образцов – 26 (из них в Западной Европе 10), гаплогруппы R1b – 118 (из них в Западной Европе 57).
Если рассмотреть все 60 балканских образцов из списка, то в них распределение гаплогрупп следующее:
G2a — 15
I2a — 13
R1b — 10
C — 5
I — 4
I2 — 2
R — 2
T1a — 2
CT — 2
E1b — 1
R1a — 1
R1 — 1
H2 — 1
G2 — 1
J2b — 1
Во всех списках верно одно – если гаплогруппу нашли, то она там была, и то если не ошиблись с типированием. Но если не нашли – то это не означает, что ее там не было, например, в соседнем захоронении. Отсутствие доказательств не есть доказательство отсутствия. Поэтому правило простое – обсуждать можно только то, что нашли, но то, что не нашли – можно только констатировать, что пока не нашли. Если, конечно, нет дополнительных и независимых данных, что то, что не нашли, там и быть не может.
Возвращаемся к сюрпризам в отношении ископаемых ДНК. Один сюрприз, о котором было упомянуто в предшествующих частях – это что в культуре ямочно-гребенчатой керамики в Эстонии нашли R1a, хотя и единичный. Хотя не совсем и единичный – по соседству, в Карелии, нашли опять R1a. Датировки – 5900-3800 лет назад и 7500-7000 лет назад, соответственно. Хотя обобщения делать преждевременно, но ясно, что R1a в те древние времена там были. А вот гаплогруппу N, которую попгенетики там предрекали, пока не нашли. Есть хорошая вероятность, что в те времена и не найдут, потому что по данным ДНК-генеалогии предки современных носителей гаплогруппы N1a1 подошли к Балтийскому морю не ранее 2500 лет назад на территории современных Эстонии-Латвии-Литвы, и не ранее 2000-1500 лет назад на территории современной Финляндии. Конечно, не исключены и более ранние заходы тех, от кого современных потомков не осталось, чьи ДНК-линии терминировались, но таких, как видно, пока не нашли.
Еще сюрприз – описанная в предыдущих частях находка двух образцов гаплогруппы R1b одной группой исследователей в нарвской археологической культуре, опять же в Прибалтике, в Латвии, и двух образцов I2a другой группой исследователей в той же нарвской культуре, но уже в Литве. Дилетанты дружно зашумели, что, мол, это показывает, что носители двух этих гаплогрупп жили вместе в одной культуре, и что положение о раздельных, как правило, миграциях в древности носителей разных гаплогрупп является неверным. Но они не обратили внимания, что и география различается, и датировки указанных образцов могут быть различными, поскольку датировки весьма широкие – R1b в нарвской культуре в Латвии датируются 7000-5000 лет назад, а I2a в Литве датируются 7500-4900 и 6440-6240 лет назад. Поэтому вполне возможно, что племена R1b и I2a жили в Прибалтике в разных местах и в разные времена. Это более вероятно, учитывая, что другие ископаемые I2a обнаружены в Западной Европе, с датировками примерно 5000 лет назад, а другие ископаемые R1b обнаружены в степных хвалынской и ямной культурах, с датировками 7215-4650 лет назад. Кстати, по данным историков (например, А.И. Юдин, 2012), насельники хвалынской археологической культуры расселялись вверх по Волге. Возможно, этим путем и далее на север они и попали в Прибалтику.
Там, где образцы более многочисленны, они показывают, что древние культуры действительно были практически однородны – двадцать G2a в одной из западно-европейских культур с датировкой 5000 лет назад, десять R1a в андроновской культуре, цепочка культур R1a от срубной до синташтинской, и далее до тагарской, таштыкской, карасукской, почти все тестированные носители культуры шнуровой керамики, имевшие гаплогруппу R1a. Естественно, в захоронениях могут оказаться включения инородных гаплогрупп – пленные, рабы, заезжие торговцы, и просто отдельные люди, принятые в племя по разным причинам, так что делать категоричные выводы «по понятиям» – это опять удел дилетантов. Надо изучать каждый случай, если, конечно, стоит соответствующая задача, и при этом формулировать конкретные и осмысленные вопросы. Но уже ясно, что с течением времени, при переходе от древнейших культур, которые были более однородными в отношении их Y-хромосомных гаплогрупп, к более недавним временам, наблюдается прогрессивное смешивание племен, с образованием полигаплогруппных. Возможно, это было следствием возрастания торгово-обменных связей, совместной защиты от неприятеля, в общем, это задача для историков.
Так, в предыдущей части этой статьи было показано, что на современных территориях Венгрии, Германиии и Испании найдено множество ископаемых ДНК с Y-хромосомными гаплогруппами CT, C, C1, C1a, F, G, G2, G2a, H, H1, H2, IJK, I, I2a, I2c, J2a, R, R1, R1b, T1a, с совокупными датировками 8000-4200 лет назад. Но делать скороспелый вывод о полигаплогруппности тех культур не стоит, потому что найденные образцы относятся к разным территориями и к интервалу времени в четыре тысячи лет. Если кто-то захочет исследовать гипотезу о «полигаплогруппности», то необходимо рассортировать все эти данные по отдельным захоронениям, датировкам и территориям, причем датировки потребуются по возможности точно (с точностью до одного-двух поколений) для конкретных захоронений. Проблема в том, что часто две гаплогруппы даже в одном захоронении вовсе не означают, что соответствующих носителей хоронили вместе, в одно время. Они могут различаться по времени веками, а то и тысячелетиями, и относиться к подзахоронениям, умышленным или случайным. Поэтому этот исследовательский проект будет чрезвычайно трудным и сложным. Даже одни и те же названия культур зачастую относятся к разным территориям, например, завершение предыдущей части статьи и начало настоящей части относится к северобалканской культуре Винча (юго-западная Венгрия), в отличие от культуры (или ее части) Винча в пригороде Белграда, Сербия. Там, в венгерской Винча, нашли гаплогруппы H2 и G2a, но в сербской Винча пока ископаемых ДНК не извлекали.
Наконец, еще один сюрприз по предыдущим частям статьи представляется в том, что в длинном перечне гаплогрупп в предшествующем параграфе нет гаплогруппы R1a. Дилетант тут же заголосит (и уже заголосили на разных форумах), что, мол, все теории на этот счет были неверны, не было в Европе гаплогруппы R1a, во всяком случае от 8000 до 4200 лет назад. Но дилетант на то и дилетант, что не умеет думать и анализировать. Он не знает, или забыл, или знать не хочет, что на указанных территориях (особенно в Германии) и в указанный период времени обнаружено на самом деле много ископаемых R1a, начиная с самой первой, в публикации Haak с сотр. (2008), в которой нашли целую группу R1a в селении Эулау с археологической датировкой 4600 лет назад. Помимо того, в Германии нашли несколько образцов R1a-M420 (датировки 4515-4065 лет назад), R1a-M198 (4515-4065 лет назад), R1a-M417 (4844-4480 и 4488-4369 лет назад), в Дании – R1a-M417 (4866-4507 лет назад), в Швеции R1a-M459 (4636-4487 лет назад), и серию более поздних образцов R1a, например, R1a-Z280 в Германии (3128-3036 лет назад). Почему в длинном перечне гаплогрупп в предыдущем абзаце нет R1a – да по той же причине, остались в соседних захоронениях, не копнули там, где они были. Повторяю – отсутствие доказательств не есть доказательство отсутствия.
Продолжим расмотрение гаплогрупп-субкладов ископаемых ДНК, результаты которых опубликованы в этом, 2017 году.
4. Северобалканская культура винча (юго-западная Венгрия), ранний каменный век, датировки 7400-7100 лет назад. Четыре образца ископаемых ДНК, два мужских и два женских, мужские имели гаплогруппы H2 и G2a.
H2-P96 (такой же, как и в культуре старчево)
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a > G2a2b2a1 > G2a2b2a1a-PF3346
Субклад PF3346 приведен на диаграмме субкладов гаплогруппы G2a ниже.
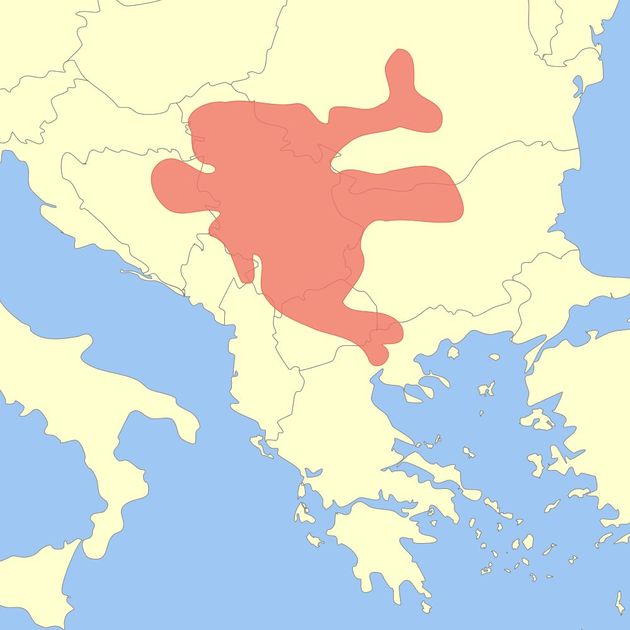

Диаграмма субкладов гаплогруппы G2a. Сиреневым цветом отмечен субклад P15, найденный в захоронении в культуре Тиссы. Такой же субклад найден в некрополе на юге Франции с датировкой 5000 лет назад. Субклады M201 (первый сверху), Р15 и PF3148 найдены в захоронениях в культурах трансдунайского позднего неолита (см. ниже).
Мы видим, что гаплогруппа H2-P96 найдена уже в трех захоронениях – дважды в культуре старчево (с датировкой 7800-7500 лет назад), и в культуре винча (с датировкой 7400-7000 лет назад). Это показывает, что данная гаплогруппа была распространена в «Старой Европе» (ранее 4500 лет назад), и погибла вместе с ней, наряду с большей частью гаплогрупп E1b, F, G2a, K, I1, I2a, R1a. В настоящее время гаплогруппы H в Европе практически нет, и в известном списке Eupedia она вообще отсутствует среди европейских гаплогрупп (ссылка).
5. Культура Тиссы на юго-востоке Венгрии, датировки 7000-6500 лет назад. Найдены пять образцов ископаемых ДНК, три мужских и два женских, мужские имели гаплогруппы G2a-P15 (отмечена сиреневым цветом на диаграмме субкладов гаплогруппы G2a, I (x I1), как в культуре линейно-ленточной керамики, см. предшествующую часть статьи, и I2a1-P37.2 (см. диаграмму субкладов гаплогруппы I2).
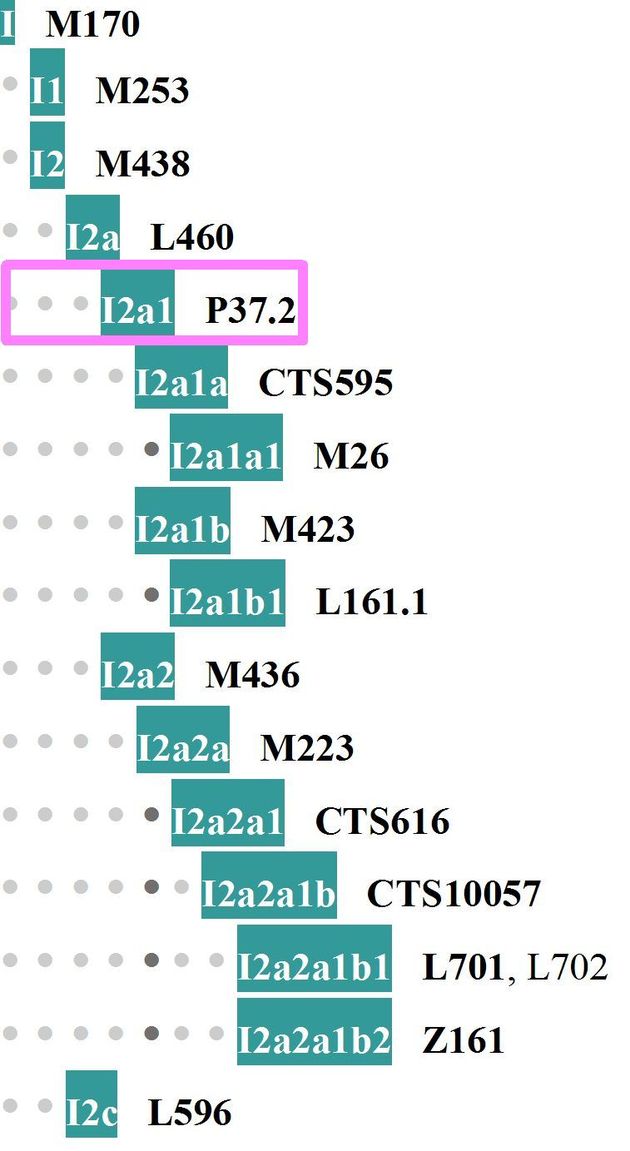
Диаграмма субкладов гаплогруппы I, на которой отмечен субклад P37.2, найденный в захоронении в культуре Тисса. На следующей диаграмме показаны нижестоящие от P37.2 субклады.
Обратим внимание на то, что субклад I2a1-P37.2 – это тот, от которого впоследствии пошли славянские линии гаплогруппы I2a (после почти полного уничтожения носителей гаплогруппы I2a в Старой Европе, и прохождения бутылочного горлышка популяции примерно 2200 лет назад, в конце прошлой эры, см. диаграмму ниже). В настоящее время эти славянские ДНК-линии являются основными по численности в некоторых странах бывшей Югославии – 71% среди боснийских хорват, 54% среди боснийцев, 37% среди хорват, 34% среди сербов, 30% в Черногории, 21% в Словении. В России, Украине, Белоруссии число носителей гаплогрупы I2a варьируется между 10 и 20%, в зависимости от региона, в Польше падает до 6%.
Ясно, что P37.2 – это далекий предковый субклад по отношению к южнославянским субкладам (см. подпись к диаграмме), он образовался примерно 21300 лет назад. Его нижеследующий субклад I2a-L147.2/CTS10228, обозначенный сиреневым цветом, наиболее распространен в гаплогруппе I2 в Восточной Европе – от Греции до Прибалтики, включая Россию, Украину, Белоруссию, страны бывшей Югославии. Он образовался примерно 5300 лет назад, и разошелся между Балканами (дунайские славяне, в частности) и Британскими островами четырьмя своими основными ветвями – S17250, Y4460, Z1785 и A2512, все четыре образовались примерно 2200 лет назад. Как правило, у их носителей имеются нижестоящие субклады. Первые три – преимущественно славянские субклады, четвертый, А2512, относительно малочисленный и встречается преимущественно у евреев.

Диаграмма субкладов, нижестоящих от P37.2. Справа – датировки возникновения субкладов, рассчитанные по снип-мутациям (группа YFull). Субклад L147.2 (отмечен сиреневым цветом) был почти полностью уничтожен в ходе гибели Старой Европы (III тыс. до н.э.), и вышел из «бутылочного горлышка популяции» примерно 2200 лет назад в виде четырех параллельных субкладов, первые три (сверху вниз, начиная с S17250) можно назвать «южно-славянскими» по преобладающим языкам и территориям, последний – субклад преимущественно евреев.
6. Трансдунайский поздний неолит (культуры лендьел и сопот), датировки 7200-6500 лет назад. Найдены 14 образцов ископаемых ДНК, 12 мужских и два женских, мужские имели гаплогруппы С (x C2), G-M201 и два G2a, F (х I), два Н-L901, H1a2a-Z14050, два образца гаплогруппы I – один был опубликован ранее, другой I (x I1, I2a2), I2 (x I2a2), а также J2a-M410, E1b-L618. Как видно, в этих культурах на протяжении примерно 700 лет наблюдается большое разнообразие Y-хромосомных гаплогрупп. Cубклады G2a в тех случаях, в которых были определены, следующие:
G2a-P15
G2a-P15 > G2a2 > G2a2a > G2a2a1-PF3148 (датировка образца 6800-6500 лет назад).
Последний субклад был также найден в культуре трансдунайской линейно-ленточной керамики (датировка образца 7200-7000 лет назад).
Комментарии:
(a) в статье использовали устаревший индекс H1b1-Z14050, но этого субклада в номенклатуре ISOGG 2017 года нет, есть H1a2a-Z14050.
(b) в опубликованном наборе ископаемых ДНК здесь впервые встречается гаплогруппа J2a. Обычно принимается, что эта гаплогруппа происходит из Месопотамии и далее распространилась в юго-восточную Европу. Древняя находка J2a в трансдунайском позднем неолите (образец датирован 6800-6500 лет назад) хотя это напрямую не подтверждает, но и не противоречит. В настоящее время гаплогруппа J2a особенно распространена в Анатолии (24%), на островах Средиземного моря (23-37%), в Греции и в центральной Италии (по 23%), в Албании (20%). В России этой гаплогруппы 3%, на Украине – 4.5%, в Белоруссии и Польше по 2.5%.
(c) Особый интерес представляет субклад E1b-L618, место которого в диаграмме субкладов гаплогруппы E отмечено желтым цветом. Последним стоит его дочерний субклад E1b-V13, который почти полностью погиб при разрушении Старой Европы, и возродился, то есть прошел бутылочное горлышко популяции 3450±350 лет назад (А.А. Клёсов, публикации 2010-2017 гг.).

7. Халколитические (медного века) культуры древней территории Венгрии, датировки 6500-5900 лет назад. Найдены 10 образцов ископаемых ДНК, 6 мужских и 4 женских, мужские имели гаплогруппы два СТ-M168 (как и в культуре линейно-ленточной керамики), два G2a, два I2a. Cубклады G2a и I2a в тех случаях, в которых были определены, следующие (их положение показано на соответствующих диаграммах выше):
G2a-P15 > G2a2 > G2a2b-L30
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a > G2a2b2a1 > G2a2b2a1a > G2a2b2a1a1 > G2a2b2a1a1c > G2a2b2a1a1c1 > G2a2b2a1a1c1a-CTS5990
I2a-L460
I2-M438 > I2a > I2a2 > I2a2a > I2a2a1 > I2a2a1b-CTS10057,
датировка образца 6500-6000 лет назад. Этот субклад был также найден в культуре линейно-ленточной керамики (датировка образца 7200-6900 лет назад).
Как мы обсуждали выше, все эти линии (G2a и I2a) почти полностью терминировались при гибели Старой Европы между 5000 и 4500 лет назад.
8. Средне-халколитический переходный период смешанных стилей керамики, под названием «протоболераз», датировки ископаемых ДНК 5900-5600 лет назад. Найдены четыре образца, три мужских и один женский, мужские имели гаплогруппы – одна G2a и две I2c (с одинаковыми субкладами), их положение показано на диаграммах выше:
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a-P303 (отмечен желтой полосой на диаграмме субкладов гаплогруппы G2a)
I2-M438 > I2c-L596
Как видим, среди ископаемых ДНК в центральной Европе появляется I2c. Это в настоящее время довольно редкая гаплогруппа, она отсутствует в списке основных европейских гаплогрупп (ссылка), и в проектах FTDNA, в которых числятся 6000 носителей гаплогруппы I1, 2000 носителей гаплогруппы I2a, есть только 300 носителей гаплогруппы I2c. Как обычно, у всех четырех ископаемых ДНК в данной серии все четыре мтДНК разные – H, J1c, N1a1a1a3, U5a1c1.
9. Баденская культура на территории современной Венгрии (в целом она расположена на территориях Чехии, Словакии, Венгрии и Австрии, см. карту), датировки ископаемых ДНК 5600-4850 лет назад. Найдены тринадцать образцов, девять мужских и четыре женских, мужские имели гаплогруппы – пять G2a, I-M170 и три I2a (все они показаны на соответствующих диаграммах субкладов выше).

G2a-P15 > G2a2 > G2a2a-PF3147.
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a-P303 (отмечен желтой полосой на диаграмме субкладов гаплогруппы G2a)
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a > G2a2b2a1 > G2a2b2a1a-PF3346
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a > G2a2b2a1 > G2a2b2a1a > G2a2b2a1a1 > G2a2b2a1a1b > G2a2b2a1a1b1-Z1815
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2a > G2a2b2a1 > G2a2b2a1a > G2a2b2a1a1 > G2a2b2a1a1c > G2a2b2a1a1c1 > G2a2b2a1a1c1a-CTS5990
I-M170
I-M170 > I2-M438
I2-M438 > I2a > I2a1-P37.2
I2-M438 > I2a > I2a1-P37.2 > I2a1a > I2a1a1-M26
10. Культура линейно-ленточной керамики на територии современной Германии, найдены 29 образцов ископаемых ДНК, из них 10 – мужских костяков, с идентифицированными гаплогруппами-субкладами, из которых пять опубликованы ранее, и пять описаны впервые в рассматриваемой работе. Датировки – 7500-7100 лет назад, то есть практически те же, что и линейно-ленточной керамики в Венгрии. Но набор обнаруженных гаплогрупп резко различается – если среди 11 образцов в той же культуре в Венгрии была только одна гаплогруппа G2a (субклад P303), то она обнаружена в 8 из 10 образцов в Германии, и среди них не было субклада Р303. Вместо линии G2a2b-L30 (как в Венгрии) там была почти исключительно линия G2a2a. Иначе говоря, на фрагменте диаграммы субкладов гаплогруппы G это субклады PF3147 и PF3148, размещенные между сиреневой и желтой полосой.
Это говорит о том, что в культуре линейно-ленточной керамики в Венгрии и в Германии в одни и те же времена жили разные племена, которые разошлись от субклада G2a2-L1259, образовавшегося 18200±1600 лет назад. Похоже, что они прибыли в Европу раздельными миграциями, и принадлежность их к одной культуре можно объяснить «культурной диффузией». Конечно, это пока основывается на небольшом наборе ископаемых ДНК.
Три другие Y-хромосомные гаплогруппы, помимо G2a, были G2-P287 (см. диаграмму субкладов гаплогруппы G), СТ и T1a, две последние были опубликованы ранее, и потому не анализировались в рассматриваемой статье. Семь гаплогрупп G2a имели следующие субклады:
G2a-P15 > G2a2 > G2a2a-PF3147 (три образца)
G2a-P15 > G2a2 > G2a2a > G2a2a1-PF3148 (четыре образца)
11. Средний неолит, территория современной Германии, археологические датировки 6600-5000 лет назад. Найдены семь образцов ископаемых ДНК, из них четыре мужских и три женских, Y-хромосомные образцы гаплогрупп I, R, G2a-PF3148, и IJK (x J), из них три первых уже были опубликованы. Образец с гаплогруппой R датирован 5590±64 лет назад, что заметно раньше прибытия в Европу (на Пиренейский полуостров) основной миграции носителей гаплогруппы R1b. К сожалению, гаплотипов в статье нет, поэтому выяснить ДНК-генеалогическое происхождение этого образца пока не представляется возможным. Он имел мтДНК HV, что, впрочем, тоже мало о чем говорит, и такие же мтДНК найдены в захоронениях культуры линейно-ленточной керамики в восточной Венгрии со значительно более древней датировкой (7500-7300 лет назад) и в баденской культуре в Венгрии с заметно более поздней датировкой (5300-4850 лет назад).
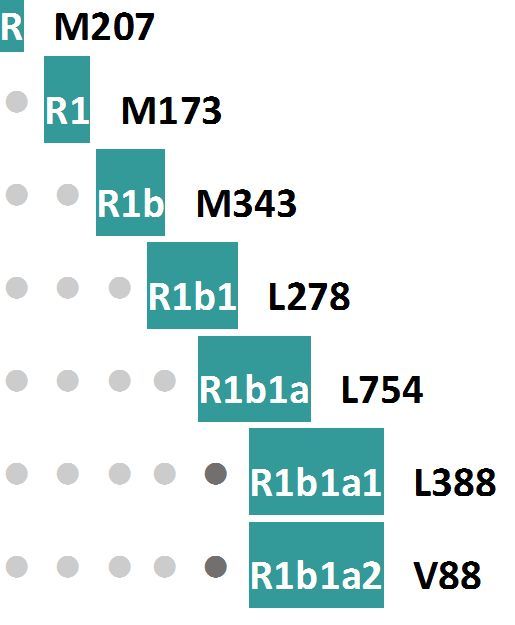
12. «Лиственная пещера» в западной части Германии, датировка 6000-5000 лет назад. Найдены четыре образца ископаемой ДНК, из них три мужских. Гаплогруппы I2a1-P37.2 (см. диаграмму выше), R1-M173 и R1b1-L278 (диаграмма ниже).
Гаплогруппа I1a1-P37.2 нам уже знакома, эта линия после ее почти полного уничтожения в ходе расселения эрбинов, носителей гаплогруппы R1b по Европе, привела к настоящему времени к появлению десятков миллионов славян Восточной Европы, от Адриатики до Балтики. Две гаплогруппы линии R1, обнаруженные в ископаемых ДНК «лиственной пещеры» (Blätterhöhle Cave), представляют древнейшие гаплогруппы, которые образовались 22000-20000 лет назад. В мезолите центральной Европы их найдено не было, но они были найдены в Прибалтике, в нарвской культуре, и определенно пришли в Европу не вместе с основной группой мигрантов на Пиренейский полуостров, а, видимо, из той самой нарвской культуры, что описано в первых разделах этой статьи; более того, субклад в нарвской культуре и в «лиственной пещере» одинаковый, R1b-L278 (образовалась 20400±3000 лет назад, https://www.yfull.com/tree/R1b/).
В «лиственной пещере», которая была открыта в 2004 году, нашли скелетные останки более 400 человек. Генетический анализ показал, что часть из них питалась в основном рыбой, другая часть – зерновыми продуктами. К удивлению исследователей, оказалось, что эти части популяции генетически не смешивались. Правда, генетики заключили, что эти популяции сосуществовали в пещере только 400-600 лет (видимо, основываясь на численности захоронений в пещере), но генетики не привели датировок, когда это было. Ископаемые ДНК в рассматриваемой статье имеют датировку 6000-5000 лет назад, так что если генетики правы в своих оценках продолжительности заселения пещеры, то не исключено, что население пещеры было изгнано или уничтожено как раз во время гибели «Старой Европы», в III тыс. до н.э. Но то, что две описанные выше популяции генетически не смешивались, живя практически в одном месте веками, в очередной раз подтверждает гипотезу, приведенную выше, что древние племена обычно не смешивались.
13. Последняя группа гаплогрупп-субкладов в статье – это иберийские (то есть пиренейские) ископаемые ДНК раннего и среднего неолита (то есть каменного века), и халколитического периода (медного века). Рассмотрим их по частям.
Ранний неолит. Датировки ископаемых гаплотипов 7500-7300 лет назад. Найдены семь образцов ископаемой ДНК, из них два мужских – гаплогруппы I2a-L161.1 и R1-V88. Оба Y-хромосомных образца были ранее опубликованы, поэтому в рассматриваемой статье их субклады не анализировали, ограничились ссылками без приведения снипов. Они нами показаны, основываясь на буквенных индексах субкладов, принимая их за индексы 2017 года, как это было по всей рассматриваемой статье. Если это индексы, скажем, 2015 года или ранее, то соответствующие снипы могут быть другими, например, R1b-V88 станет R1b-M269. Расположение обоих субкладов на диаграммах соответствующих субкладов гаплогрупп R и I приведено выше.
Средний неолит. Датировки ископаемых гаплотипов 5900-5600 лет назад. Найдены четыре образца ископаемой ДНК, из них два мужских – гаплогруппы Н2 и I2a-Z161. Оба Y-хромосомных образца были ранее опубликованы, поэтому в рассматриваемой статье их субклады не анализировали, ограничились ссылками без приведения снипов. Снип для I2a нами реконструирован по виду субклада.
Халколит, датировки ископаемых гаплотипов 5600-4350 лет назад. Найдено 26 образцов ископаемой ДНК, из них 13 мужских – гаплогруппы I (три образца), I2a (восемь образцов), G2a (2 образца). Половина Y-хромосомных образцов были ранее опубликованы, поэтому в рассматриваемой статье их субклады не анализировали, ограничились ссылками без приведения снипов. Cубклады G2a и I2a в тех случаях, в которых были определены, следующие (их положение показано на соответствующих диаграммах выше):
G2a-P15 > G2a2 > G2a2a-PF3147.
G2a-P15 > G2a2 > G2a2b > G2a2b2 > G2a2b2b-PF3359.
I2-M438 > I2a > I2a2-M436 (два образца).
I2-M438 > I2a > I2a2 > I2a2a-M223 (пять образцов).
I2-M438 > I2a > I2a1-P37.2 > I2a1a > I2a1a1-M26
Подводя итоги, повторим вслед за авторами обсуждаемой статьи, что в период неолита-халколита (8000-4850 лет назад) на территории современной Венгрии наиболее распространенной гаплогруппой Y-хромосом (найденной среди ископаемых ДНК) была гаплогруппа G (в основном G2a с нисходящими субкладами, линии G2a2a и G2a2b), на которую приходится примерно 40% от всех находок. Эти две линии (со снипами PF3147 и L30, соответственно) расходятся в указанном периоде очень далеко, и могут относиться или к разным племенам, которые вместе мигрировали в Европу, или к одному, но очень древнему племени. То, что эти далеко расходящиеся ветви принадлежали к одним и тем же археологическим культурам (порой в их динамике), можно считать указанием, что между ними происходил вполне интенсивный культурный обмен, или что они перенимали технологии друг у друга тем или иным способом.
Второй наиболее распространенной гаплогруппой, найденной в ископаемых ДНК, была гаплогруппа I (в основном I2a с нисходящими субкладами, линии I2a1 и I2a2), на которую приходится примерно 32% от ископаемых гаплогрупп всех находок. Эти две линии (со снипами P37.2 и M436, соответственно) тоже расходятся далеко в указанном периоде, и относятся к тем же археологическим культурам, что и гаплогруппы G2a. Это означает, что культурный (и вещевой) обмен происходил «поперек» родовых, наследственных образований. Видимо, оседлый образ жизни располагал к культурным диффузиям, в отличие, как представляется, от кочевого образа жизни. В те же времена, или несколько позже, в центральную Европу приходят носители гаплогруппы R (захоронение с датировкой 5654-5527 лет назад), R1 (датировка 5337-5034 лет назад) и R1b (5958-5344 лет назад), а на Пиренеях обнаружена ископаемая гаплогруппа R1b-V88 (7294-7066 лет назад). Это все исключительно древние субклады, как и древние датировки захоронений, и их носители не сопровождали носителей гаплогруппы R1b-M269-L151 (и их нижестоящих субкладов U106 и P312, образовавших культуру колоколовидных кубков с последующим заселением Европы). Похоже, что древние гаплогруппы R1b могли быть принесены в Европу из нарвской культуры Балтики (обнаруженные там захоронения с гаплогруппой R1b датируются 7690±100 и 7030±225 лет назад), то есть ранее центрально-европейских. В нарвскую же культуру эрбины скорее всего прибыли из приволжских степей (хвалынская культура и предшествующие ей среднеазиатские миграции, из Сибири через южный Урал и северный Казахстан).
Маршруты прибытия в Европу носителей гаплогруппы G2a пока твердо не установлены, но есть данные, позволяющие полагать, что они прибыли туда вскоре после ухода ледника, со стороны Передней Азии, где датировки общих предков этой гаплогруппы уходят во времена 14 тысяч лет назад. Возможно, прибытие этой гаплогруппы через Анатолию на Балканы и воспринимается генетиками как прибытие «анатолийских и левантийских фермеров». Гаплогруппа I (и ее дочерние I1 и I2a) образовались 43000±3000 и 27500±2800 лет назад, соответственно, и могут знаменовать собой кроманьонские времена в Европе. По каким-то, пока неизвестным причинам, гаплогруппа I1 встречается в захоронениях древней Европы значительно реже, чем I2 (I2a), но в итоге все перечисленные гаплогруппы Старой Европы, кроме R1b, почти полностью погибли в III тыс до н.э.
Фактом является то, что ни в одном случае из 82 ископаемых ДНК в Венгрии, Германии и Испании (c датировками от 7800 до 4350 лет назад) ни разу не встретилась гаплогруппа R1a, хотя она намного раньше встречается в Прибалтике (культура ямочно-гребенчатой керамики, другие мезолитические культуры), и была найдена в Германии (Эулау) с археологической датировкой 4600 лет назад. Либо не там искали, либо пути ее миграции в Европу нам еще недостаточно известны. Ясно, что примерно 5500 лет назад (гаплогруппа R1a-Z645) она в Европе была, там же началось расхождение ее пра-индоевропейского языка на ветви, и там же примерно 5200 лет назад возникла культура шнуровой керамики, без сомнения, культура носителей R1a, скоре всего R1a-Z645 > Z283 > Z283 > Z280.
Так что, как уже обсуждалось выше, мы наблюдаем явное противоречие, или несогласованность в экспериментальных данных, и нас, вполне возможно, ожидает лавина данных по наличию гаплогруппы R1a в древней Европе. Возможно, на Балканах и/или в трипольской культуре.
История обнаружения гаплогрупп Y-хромосомы у степняков следующая. В 2015 (Mathieson и др., Nature) было сообщено о первой находке ископаемой скифской ДНК, с гаплогруппой R1a-Z93-Z2123 (датировка 2305±90 лет назад), то есть конец прошлой эры, классические скифские времена. Находка была сделана в волжских степях Самарской области. Следующие два образца были извлечены из двух хазарских захоронений на нижнем Дону (VIII-IX вв н.э.), что технически позднее скифских времен, но может дать свидетельство о гаплогруппах их возможных потомков, тоже степняков. Обе гаплогруппы оказались R1a-Z93 (Klyosov, Faleeva, Adv. Anthropol., 2017). В марте 2017 года опубликована статья (Unterländer и др., всего 25 авторов, среди которых опять David Reich из отдела генетики Гарвардской медицинской школы), в которой было сообщено об идентификации четырех гаплогрупп скифов, двух R1a (Z645 и Z645-Z93), одной R1b-Z2123 и одной Q1a-F903. R1a-Z645-Z93 была из пазырыкской культуры Алтая, R1a-Z645 из тувинской Алды-Бель, R1b-Z2103 «ранняя сарматская», к юго-западу от Урала, Q1a – из восточного Казахстана (см. карту).
Собственно, на этом данную информацию можно было бы закончить, потому что гаплогруппы в статье не обсуждались, и даже не приводились (им было уделен десяток строк в приложении объемом 92 страницы, с описанием сугубо технических деталей типирования). Так, было сообщено, что R1a-Z645 скорее всего недотипирован. Что же тогда обсуждалось? А то, что я писал выше о «геномных» статьях Reich – вязко, мутно, неопределенно, противоречиво, и в итоге… что оказалось в итоге, опишу и прокомментирую ниже.
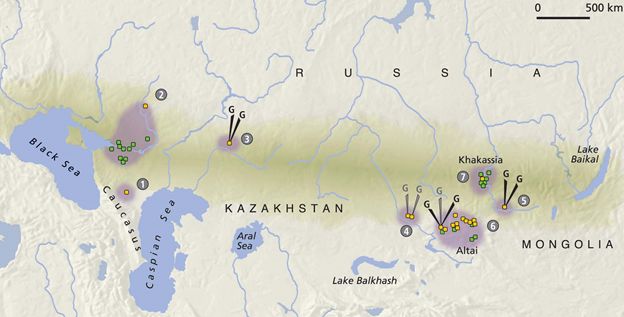
География ископаемых ДНК скифов (по определению в рассматриваемой статье). Образцы с 1 по 3 в статье называют «западными скифами», с 4 по 7 – «восточными». Y-хромосомные гаплогруппы определяли в образцах 3 (R1b-Z2123, «ранний сармат», V-II вв. до н.э., к юго-западу от Урала), 4 (Q1a-F903, Восточный Казахстан, IX-VII вв. до н.э.), 5 (R1a-Z645, культура Алды-Бель, VII-VI вв. до н.э., Тува, Аржан-2), 6 (R1a-Z93, пазырыкская культура, IV-III вв. до н.э., Алтай). В участках 1, 2, 7 Y-хромосомные гаплогруппы не определяли, только мтДНК и геномные данные.
По статье красной нитью проходит, начиная с Абстракта, что геном показал, что скифы «могут быть наилучшим образом описаны как представляющие собой смесь компонента ямной культуры и восточно-азиатского компонента». На самом деле, как показывают найденные гаплогруппы скифов, среди них есть R1b-Z2123, и R1a и Q1a. Действительно, первый субклад был во множестве найден в ямной культуре, а последний – типичный для сибирских народов. Фокус в том, что «широкогеномные» всё смешивают и усредняют. Вот и получается «смесь компонентов». На деле – это разные рода, которые поколение за поколением наследуются по мужской линии. Прочитаем еще раз выделенное курсивом – и что это дает для понимания скифов? Чем это количественно отличается от «да, Скифы мы! Да, азиаты мы»? Вообще получение данных и их обсуждение на уровне «смесь компонентов» и «похожести», «admixtures» и прочее – это шаги назад по сравнению с подходами ДНК-генеалогии.
Но дело еще хуже – ниже показана диаграмма «геномных компонент» для ряда выборок, составленных из ископаемых ДНК разных культур и времен, и представленная в рассматриваемой статье. Мы видим, что все они практически одинаковы, не считая минорных вариантов. Рядом сидят ископаемые ДНК из культур потаповской и полтавкинской, первая культура R1a, вторая – R1b. Они практически идентичны как по «синей» компоненте, так и по «зеленой», но что означает та и другая – «широкогеномные» не знают.
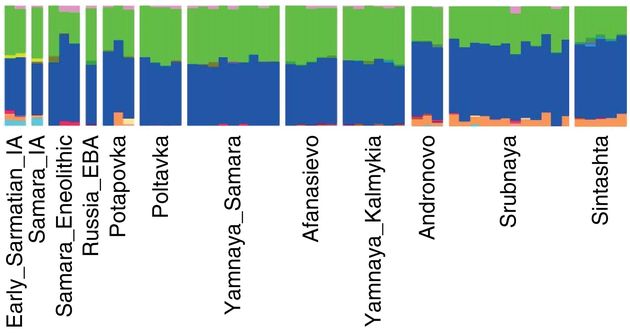
Проблема усугубляется тем, что сплошную синюю компоненту имеют ископаемые ДНК из Моталы (Швеция), все пять из которых имели гаплогруппу I (одна I, три I2a и одна I2c с датировками 3780±130 лет назад для первой, и 7730±180 лет назад для остальных четырех), и такая же синяя – ископаемые WHG (то есть «западные охотники-собиратели»), которые имели в большинстве гаплогруппу G2a, и на втором месте I2a, в сумме примерно 70% от всех. Так что означает синий цвет, который занимает примерно 50% «компонент» и в ямной культуре (R1b), и в полтавкинской (R1a), и в потаповской (R1a), и в андроновской (R1a), и в срубной (R1a), и в синташтинской (R1a), и в афанасьевской, которую авторы на основании «сходства с ямной» зачисляют в R1b, причем считают, что те R1b пришли в афанасьевскую культуру именно из ямной культуры, а не в обратную сторону?
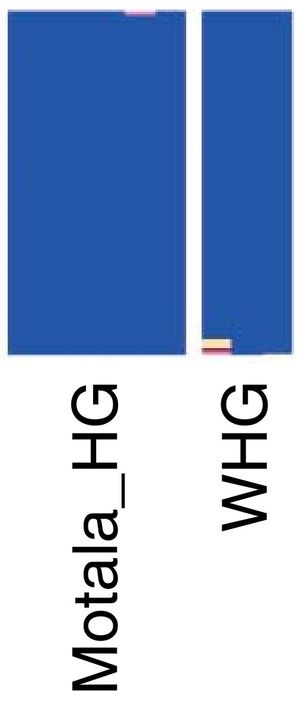
Тот же синий цвет – и у ранних сармат, у которых найдена гаплогруппа R1b-Z2103. То, что сарматы имеют ту же гаплогруппу, что и ямники, и цвет почти одинаков (кроме дополнительных минорных цветовых «компонент» у сармат) – замечательно, но что делать с тем, что такие же цвета и у культур с гаплогруппой R1a?
«Широкогеномные» и прочие попгенетики, конечно, скажут, что, мол, причем здесь гаплогруппы Y-хромосомы, цвета другие компоненты ДНК определяют. Встречный вопрос – какие? Да что бы ни определяло, сравнение (и интерпретации) ведь по цветам проводятся. Безотносительны здесь и мтДНК, так как у образцов из Моталы (Швеция) они все U (U2 и U5) и одна T1a1, а у «раннего сармата», например, мтДНК М. А у срубной культуры – букет из U5, H5, J2, T2, H3. Так что, друзья, измеряете? Или то, что компьютер покажет, после разноплановых сортировок?
Иначе говоря, если цвета что-то и показывают, то за пределами тех интерпретаций, которые «широкогеномными» делаются. Вот и получаются «выводы», что «наш анализ показывает, что восточные скифы (напомним, что это скифы гаплогрупп R1a и Q1a – AAK) образовались наиболее вероятно из одной единственной популяции, в то время как два других сценария, которые предполагают, что восточные скифы разошлись из других ранее разошедшихся популяций, получил очень малую степень статистической поддержки». Замечательно, не так ли? R1a и Q1a – имеют одно и то же происхождение. Так показал компьютер. Авторы продолжают – «генетические дистанции между комбинированными (! – ААК) группами скифов, восточными и западными (напомним, что западные – это R1a и R1b, восточные – R1a и Q1a) показывают, что лучше всего подходит мультирегиональная модель, хотя модель западного единого происхождения также получает некоторую поддержку, и не может быть сброшена со счетов». Информативно, не так ли? В то время как беглый взгляд на гаплогруппы сразу показывает, что они, R1a, R1b, Q1a имеют, разумеется, различное происхождение.
Вообще пытаться найти «происхождение», манипулируя усредненными параметрами с неясным смыслом – дело совершенно проигрышное. И тут же авторы продолжают – «наши результаты показывают, что западные и восточные группы скифов возникли независимо». И это опять неверно, потому что «восточные» группы скифов у авторов – это R1a и Q1a, нельзя их объединять для «выяснения происхождения». Авторов опять подводит привычка всё усреднять. Но это грубое нарушение простейшего математического (даже арифметического) правила – нельзя усреднять разные сущности, как в данном случае популяции R1a и Q1a. Большая ошибка. А авторы опять за своё – «наше моделирование свидетельствует в пользу непрерывного потока генов между скифскими группам железного века, указывая на асимметричный поток генов от западных к восточным группам, скорее, чем в обратном направлении». Очень информативно, не так ли? Хочется спросить – вам что, делать нечего? Вы не хотите даже на гаплогруппы посмотреть, не говоря о гаплотипах, поэтому наводите седьмую воду на киселе, зачем вам в такой мутной ситуации не менее мутное «направление потока генов»? Что это вам даст, на какую историческую загадку ответит? Да при такой вязкости и мутности расчетов и изложения вообще ни на что не ответит.
Поскольку мы знаем, что в андроновской культуре обнаружена древняя ископаемая гаплогруппа R1a-Z93, и та же обнаружена в пазырыкской культуре, то понятно, что между ними есть «linkage», то есть генеалогическая связь. Но авторы опять не хотят смотреть на гаплогруппы-субклады, и вместо того используют программу ABC, рассчитывают «низкие величины FST между этими группами», и приходят к выводу «об очень сильной поддержке наличия такой связи». Ну, замечательно, нашли-таки. Хотя это и на глаз видно, без расчетов, просто смотря на R1a-Z93 там и там. Однако не тут-то было – «эти симуляции не позволяют полностью выявить картину генетического разнообразия, наблюдаемую в популяциях железного века, предполагая, что истинная демографическая история предковости популяций железного века может быть более сложная, чем рассматривалась здесь». Немудрено – когда стреляют из пушки по воробьям, всегда наблюдаются разные усложнения, не предполагаемые заранее.
Выводов в статье нет, что тоже не удивительно. Туда, в выводы, нечего и помещать. Разве что выявили четыре ископаемых гаплогруппа-субклада, но об этом в статье вообще не упомянуто, только, как уже говорилось, в нескольких строках в Приложении. В Абстракт вошел только один «вывод», который авторы, видимо, особенно ценят – «моделирование предлагает независимое происхождение западной и восточной (скифских) групп с потоком генов между ними, что вероятно (удобно) объясняет поразительную однородность их материальной культуры. Мы таже нашли свидетельства, что значительный поток генов из восточной в западную Евразию мог происходить в течение раннего железного века». Вот, собственно, и всё, чем закончилось изучение 113 образцов скифов (в основном мтДНК).
В мае этого, 2017 года вышла статья, которую многие давно ожидали. Она поднимала вопросы истории басков, или, скорее, региона, в котором позже появились баски со своим неиндоевропейским языком, столь редким в современной Европе. Статья имела два названия – одно длинное, неуклюжее и нечитаемое, второе, альтернативное, намного короче: «Популяционная геномика археологического перехода в западной Иберии», и это опять был препринт в сетевом издании bioRxiv. Это издание публикует статьи в предварительном, нерецензированном виде, хотя эти статьи читаются и обсуждаются не менее активно (и обычно даже больше), чем рецензируемые статьи в академических изданиях. Это в очередной раз обнажает лицемерие попгенетиков, в первую очередь, Балановских, которые уже много лет, пытаясь дискредитировать другие издания, которые не оставляют камня на камне от безграмотных писаний попгенетиков, объявляют, что нерецензируемые издания «ненаучны», как «ненаучны» и издания в открытом доступе. При этом умалчивают, что, например, PLOS ONE, один из передовых журналов в генетике, является журналом в открытом доступе, и, кстати, взимает 2500 долларов за каждую публикуемую там статью. Nature, кстати, тоже не публикует статьи бесплатно, и не выставляет их в открытом доступе, их либо нужно покупать постатейно, либо выписывать за деньги сам журнал, что я, кстати, и делаю уже много лет.
В статье 18 авторов из разных стран мира – Португалии, Ирландии, Швейцарии, Германии, Англии. Но, к сожалению, статья опять попгенетическая, и потому толку от нее немного, за исключением приведенных в ней гаплогрупп восьми образцов скелетных останков из разных археологических участков Португалии. Одно захоронение относится к среднему неолиту (по данным авторов, 6200-5500 лет назад), в нем найдена гаплогруппа I. Четыре захоронения – позднего неолита/халколита (медный век, он же энеолит), авторы его не датируют, но обычно это 5500-4500/4000 лет назад, и нашли там два образца I2a1b, один I2a1a, и один G2a2a1. Наконец, в двух захоронениях среднего бронзового века (по датировкам авторов 3740-3430 лет назад) нашли три гаплогруппы R1b. К сожалению, снипы авторы не привели, а привели, делая типичную ошибку попгенетиков, только названия субкладов, которые меняются каждый год и чаще, и остается только гадать, какие снипы на самом деле у тех R1b1a2, R1b1a2a1a2 и R1b1a2a1a2, которые они привели. Если названия субкладов современные, 2017 года, то первый – это R1a-V88, а двух последних в классификации вообще нет, тем более что они должны быть нижестоящими от V88, номенклатура которых крайне ограничена. В общем, нет таких. Но загадка довольно быстро решилась, авторы действительно использовали устаревшую номенклатуру, которая была действительна в 2013-2015 годах, в 2016-м она уже поменялась, и опять поменялась в 2017-м. Попгенетики действительно необучаемы.
Короче, один из R1b оказался M269, и два других P312. Всё стало на свои места. Действительно, как было описано еще в статье в Advances in Anthropology (Klyosov, 2012), носители R1b-M269-L11 (он же L151 и он же P311) прибыли примерно 4800 лет назад на Пиренеи, и от них образовались P312 и U106. Датировка общего предка для современных 829 гаплотипов R1b-U106 в 111-маркерном формате равна 3584±359 лет назад (без округления, Klyosov & Kilin, Adv. Anthropol., 2016), такая же датировка относится и к общему предку современных R1b-P312, как было показано еще в 2009 году (Klyosov, J. Gen. Geneal.) на 464 гаплотипах, и с тех пор многократно воспроизводилась. И вот теперь те же R1b-P312 найдены в захоронении в Португалии с археологической датировкой 3740-3430 лет назад, что практически совпадает с расчетами ДНК-генеалогии.
В остальном статья про «популяционную геномику западной Иберии» не слишком интересна или информативна. Все ее основные выводы уже были получены ранее в рамках ДНК-генеалогии. В частности, что не было в Иберии «популяционной непрерывности», или «палеолитической непрерывности», поскольку, как ДНК-генеалогия давно показала, около 5000 лет назад пришли эрбины и кардинально изменили там древнее, коренное население (найденные их гаплогруппы I, I2a и G2a соответствуют автохтонным гаплогруппам центральной и юго-западной Европы, уже неоднократно найденным при раскопках древних некрополей, в частности, в северной Испании и на территории Франции). В целом статья изобилует недоразумениями, вынесенными из «широкогеномного анализа», например, о вкладе «степной интрогрессии в ДНК португальцев бронзового века». Заметьте, опять эти вязкие термины, типа «интрогрессия», когда авторы не могут сказать напрямую, в простых терминах, и им надо изворачиваться, пытаясь уклончиво намекать. Не знают – так зачем эти выверты? Опять пассажи о приносе индоевропейского языка из степей в центральную Европу, одновременно с тем, что те же степи принесли в Иберию неиндоевропейский язык. При этом ни слова, чем же эти «степи» все-таки так различаются, что одни приносят в Европу ИЕ языки, другие не-ИЕ языки. Похоже, что это старая «курганная» гипотеза М. Гимбутас, доведенная до абсурда. Опять эти стандартные и неопределенные пассажи о «значительном вкладе местных охотников-собирателей в популяцию неолитической Иберии». Кто-либо знает, что всё это означает? Конкретно? А это ведь у авторов в Абстракте, значит, полагают, что особенно важная часть исследования. Кстати, опять списанные из других (ошибочных) статей попгенетиков повторы про то, что «западные линии R1b проистекают из комплексов культуры шнуровой керамики и колоколовидных кубков». Видимо, никто им не сказал, что в культуре шнуровой керамики далеко преобладающей является гаплогруппа R1a, а в культуре колоколовидных кубков – R1b.
Авторы попадаются в очередную банальную ловушку дилетантов, повторяя, что «гаплогруппа R1b проявляет сильную ассоциацию со степными миграциями». Заметьте, опять это типичное уклончиво-вязкое «проявляет ассоциацию». Интересно, когда носители R1b прибыли в Австралию и Новую Зеландию, там тоже была «ассоциация со степными миграциями»? Неужели так трудно сообразить, что миграции идут далеко необязательно по прямой, и в данноми случае они были сначала сибирскими, потом уральскими, потом среднеазиатскими, потом степными, затем кавказскими, ближневосточными, североафриканскими, через-гибралтарскими, пиренейскими и так далее, по прошествии тысяч лет и тысяч километров. Почему непременно только степными? Линейное мышление? Это – бич попгенетиков.
Завершу этот раздел тем, что читая статью про «популяционную геномику в западной Иберии», авторов становится более и более жалко. Они заполняют статью, и особенно дискуссию, набором ничего не значащих, уклончивых, вязких фраз. Последние две фразы раздела «Дискуссия», которые обычно озвучивают основные выводы статьи, в данном случае следующие: «Наблюдаемый нами определенный, но ограниченный (генетический) вклад бронзового века резонирует с неполной индо-европейской лингвистической конверсией на полуострове, хотя имеются последующие генетические изменения в Иберии, и определение горизонта для языкового сдвига пока невозможно. Это контрастирует с северной Европой, для которой отсутствуют доказательства для более ранних языков, и в которой были более масштабные миграции Бронзового века». Кто что понял? И вот так – вся статья.
Прошу понять меня правильно. «Широкогеномные» делают, наверное, полезную работу, пытаясь отработать методологию геномного анализа ископаемых ДНК. На данном этапе подходы совершенно примитивные, с массой допущений и приближений, и попгенетики пытяются замаскировать эту примитивность изощренной замысловатостью многочисленных компьютерных программ. Но изощренная замысловатость никогда и никому не помогает, хотя на какое-то время обманывает неискушенного читателя. Но поскольку ответы на вопросы они найти, как правило, не могут, да и вопросы обычно не формулируют, поэтому и возникают те ничего не значащие, уклончивые, вязкие фразы. Вместо того, чтобы написать – ну, не знаем мы, ничего не получается, или получается какая-то ерунда, типа из R1b получается R1a и наоборот, они занимаются откровенным блефом. Есть единственный путь, технически без особого труда реализуемый сейчас – это в каждом случае геномного анализа древних ДНК определять гаплотип, который обычно без труда определяется из тех же геномных данных для каждого тестированного мужчины, или определять гаплотип параллельно, обычными методами. Наличие гаплотипа сразу резко сократит число степеней свободы геномного анализа, то есть направит геномный анализ в нужном направлении. Понятно, чего опасаются «широкогеномные» попгенетики – им понадобится осваивать ДНК-генеалогию, или еще хуже – что подходы ДНК-генеалогии сделают ненужными многие подходы «широкогеномного» рассмотрения. Окажется, что результаты ДНК-генеалогии настолько ясные, что геномный анализ окажется излишним. А как же тень на плетень напускать? Как же без блефа и обмана, к которому попгенетики так привыкли? Вот и имеют они, что имеют.
В конце мая этого года вышла статья, которая по своему посылу была беспроигрышной, поскольку рассматривала ДНК древнего Египта. Это – традиционная исключительно популярная тема. Правда, авторы (16 человек по главе с Иоханнесом Краузе из Германии) умудрились дать ей столь же исключительно скучное название – «Геномы мумий древнего Египта предполагают увеличение суб-сахарской африканской предковости в пост-римском периоде». Впрочем, поскольку научные статьи мало кто читает, и народ кормится в основном примитивными пересказами популярных изданий, в основном сетевых, то у «масс» создалось представление, что изучали ДНК древних фараонов (именно с ними ассоцииируются мумии), и все на свете о них узнали. Иначе говоря, потрясающее достижение геномной науки, так сказать.
Свой вклад внесли и российские СМИ. Так, сайт RusNews в материале под названием «Анализ ДНК почти сотни египетских мумий шокировал ученых» более половины статьи посвящено ДНК фараона Тутанхамона, хотя в исходной статье в журнале Nature Communications ничего этого не было. Некоторые российские популярные пересказы вынесли Тутанхамона даже в заголовки своих статей, подавая это как сенсацию.
На самом деле ДНК ни Тутанхамона, ни других фараонов в исходной статье не изучали. Изучали ДНК 90 мумий обычных горожан (видимо, относительно состоятельных, чтобы заказать свое мумифицирование самим или через родственников) из одного египетского города под названием Абузир-эль Мелек, в ста километрах от Каира и пирамид Гизы выше по течению Нила. Раскопки там были проведены более ста лет назад, и с тех пор костные останки хранятся в двух археологических коллекциях в музеях Германии. Эти образцы относятся к трем историческим периодам древнего Египта – пре-птолемейскому (Новое царство, Третий переходный период и Поздний период, в совокупности XVI-IV вв. до н.э.), птолемейскому (IV-I вв до н.э.) и римскому (30 г. до н.э. – 395 г. н.э.). Но Y-хромосомные гаплогруппы определили всего у трех мумий, с датировками 2690±100 лет назад, 2680±100 лет назад и 2070±48 лет назад. Как видим, до времени жизни фараона Тутанхамона там далеко, он жил во времена Нового царства (18-я династия), приблизительно 3340-3359 лет назад. Приблизительно – потому что датировки у нескольких ведущих исследователей различаются в пределах 10-15 лет.
Как обычно у «широкогеномных» популяционных генетиков, гаплогруппы тех трех человек в основном тексте статьи даже не упоминаются, ну, неинтересно это попгенетикам. Тем более, не упоминаются гаплотипы, этого у «широкогеномных» попгенетиков вообще не бывает, они их не определяют. Гаплогруппы упоминаются только в приложении под номером 3, и сообщается, что они у двоих определены как J, глубже по сообщениям авторов им пройти не удалось, субклады неизвестны. У третьего – гаплогруппа E1b-V22. Первые два по датировкам «пре-птолемейские», 770-780 лет до н.э., третий – «римский период», последний век до н.э. (97 – 2 гг. до н.э.). На этом исследование авторов можно было бы и заканчивать, больше ничего интересного (на мой взгляд) там нет. Да, собственно, и это не очень интересно. Можно добавить, что энтузиасты из сети провели более глубокое исследование опубликованных геномных данных, и нашли, что первые две гаплогруппы – практически с одинаковой датировкой – J1a-Z2329 и J2b1-PF7314, третья, более недавняя – E1b-V22, как авторы и сообщили, ссылка.
Представляет некоторый интерес то, что обнаруженная у «пре-птолемейской» египетской мумии гаплогруппа J1a-Z2329 представляет собой нижестоящий субклад от J1-P58, который чаще всего среди евреев встречается у коэнов, верхнего эшелона иудейских священников, предполагаемых прямых потомков библейского первосвященника Аарона, брата Моисея. Правда, это пока навязанная еврейскими авторами версия, и те авторы не упоминают, что та же линия во множестве встречается у арабов Палестины, Саудовской Аравии, Ирака, Ливана и других арабских стран. И неудивительно, поскольку субклад J1-P58 образовался примерно 11700 лет назад, задолго до библейского Авраама, как и его нижестоящий субклад, Z2329, который образовался 5700 лет назад. Поэтому назвать соответствующий гаплотип «модальным гаплотипов коэнов» можно только при полном и умышленном игнорировании таких же арабских гаплотипов.
Далее, хотя в исходной статье этого тоже нет, современные египтяне имеют в основном те же три гаплогруппы, 21% J1, 6.5% J2 и 46% E. Поэтому ничего неожиданного в найденных гаплогруппах мумий нет. Естественно, это может быть случайным совпадением, что в трех мумиях из 90 через две с лишним тысяч лет найдены три основные гаплогруппы современного Египта, но что есть, то есть. Поэтому и неудивительно. Повторяю, что никакого отношения к гаплогруппе Тутанхамона, предположительно древней форме R1b-M269, найденные гаплогруппы не имеют – ни по датировкам, ни по социальному статусу, ни по самим гаплогруппам.
Теперь несколько слов по гаплогруппе и гаплотипу Тутанхамона, поскольку в этом отношении в статьях, научных и популярных, сплошная путаница. На самом деле там несколько простых положений.
1. Египетская группа исследователей под руководством министра по делам античности Египта Захи Хавасса провела изучение ДНК мумии фараона Тутанхамона (во всяком случае, того, кто был идентифицирован как Тутанхамон), и при изучении его Y-хромосомы получила, но не сразу идентифицировала (или не хотели обнародовать) серию типовых электрофореграм, характеризующих маркеры Y-хромосомы. Они обычно выглядят как серия пиков, каждый на месте соответствующего маркера, и высота их соответствует числу тандемных повторов в маркере. Любой специалист с опытом работы по расшифровке таких электрофореграм без труда скажет, какой там гаплотип.
2. Министр Хавасс со своими помощниками по изучению ДНК Тутанхамона выступил на телевизионной передаче по каналу «Discovery» 17 февраля 2010 года, и там помощник показал на камеру лист с электрофореграммой в сопровождение слов министра о том, что ДНК фараона расшифрована. Несколько зрителей сделали снимок с экрана, и после этого расшифровать показанный 16-маркерный гаплотип было делом нескольких минут.
3. Один из таких энтузиастов сразу выставил данные на форуме RootsWeb, но опубликованы они были несколько позже швейцарской компанией iGENEA, и воспроизведены в моей статье в журнале Advances in Anthropology (том 2, №2, стр. 87-105, 2012): 13 24 14 11 11 14 10 13 13 30 16 14 19 10 15 12. Это – один из вариантов неполного стандартного 17-маркерного гаплотипа, в котором опущен последний маркер DYS635. Первые 10 маркеров – обычный 12-маркерный гаплотип с опущенными DYS385a и DYS385b, и последующие – DYS 458, 437, 448, GATA H4, DYS 456, 438.
4. Поскольку гаплотип был получен нестандартным способом, то есть не через научную публикацию, то по правилам «академической науки», которые вполне разумные, он не был засчитан. Но беда «академической науки» в том, что она при этом даже не рассматривает возможность, что такое может быть, что гаплотип вполне может быть правильным, со всеми вытекающими из этого последствиями. Леность мысли в академической науке поразительна! Иначе говоря, в академической науке обычно не принято рассматривать альтернативные варианты, пусть в виде запасных. Поэтому так часто академическая наука, точнее, ее исполнители, заходит в глубокие тупики, выход из которых весьма болезнен. Это тоже следствие лености мысли, а если напрямую – следствие массового непрофессионализма в академической науке. Я часто называю это «лаборантским мышлением», то есть выучка работать по стандартным прописям. Бич современной науки. Крайняя степень такого непрофесионализма – это использование слов «лженаука», «псевдонаука», именно тогда, когда нежелание разобраться в вопросе становится по сути патологическим. Обратите внимание – те, кто использует слова «лженаука» или «псевдонаука», никогда не приводят конкретных обоснований своих слов. У них, как правило, для этого просто не хватает квалификации.
5. Вот и в данном случае авторы статьи про геномный анализ мумий, упомянув про ДНК Тутанхамона, специально подчеркнули, что там не было тщательных тестов на загрязнение образца теми, кто этот образец исследовал. Естественно, такие тесты нужны, но авторы не озаботились даже подумать о том, что (предполагаемый) гаплотип Тутанхамона настолько необычен, что оказаться загрязнением от исследователей практически невозможно, такая вероятность крайне мала. Как показано в публикации из Adv. Anthropology (см. выше), этот гаплотип безусловно относится к субкладу R1b-M269, но его DYS439 = 10 встречается только у 0.5% носителей гаплогруппы R1b в Европе. Такое же бездумное предположение о загрязнении образца ДНК Тутанхамона сделала автор статьи в журнале Nature (Watson, 2017), под названием «ДНК мумий раскрыла предковость древних египтян».
В той же статье в Adv. Anthropology (Klyosov, 2012) показано, что общий предок носителей гаплогруппы R1b-M269, в которую входил Тутанхамон, жил примерно 8175 лет назад. Это – времена, предшествующие хвалынской археологической культуре, в которой найден образец R1b-M269-M415 (= L278, возник примерно 20400 лет назад), с археологической датировкой 7215-6015 лет назад (6615±600 лет назад). Скорее всего, этот общий предок жил на Южном Урале, или в Северном Казахстане, в культурах Тургайского прогиба, далее его потомки прошли через хвалынскую археологическую культуру, ямную культуру, оттуда, с Русской равнины, прошли через Кавказ в Месопотамию, и далее, видимо, через Египет к Атлантическому побережью и на Пиренейский полуостров, откуда заселили Европу, в основном между 4500 и 3500 лет назад.
6. Это, конечно, не означает, что в Египет 5000 лет назад пришли «русские», русских пять тысяч лет назад не было, да еще после тысячелетий жизни тех древних мигрантов на Кавказе и в Месопотамии. Забавно, что многие комментаторы моего телевыступления на ДеньТВ восприняли эти именно так, что «русские посадили фараона в древнем Египте». На самом деле это не столько забавно, сколько печально, что те комментаторы не умеют думать и соображать. Комментаторы вообще массово играют в испорченный телефон. Но описываемые миграции действительно шли через Русскую равнину, и ископаемые ДНК в хвалынской и ямной культуре тому прямое доказательство. Только они были не русскими, а эрбинами, русских от них осталось на Русской равнине только 5%, и прямые предки их в Египет не ходили, что бы там Фоменко и Носовский с Петуховым ни говорили.
Комментарии некоторых специалистов в отношении ДНК Тутанхамона действительно забавляют. Цитата – «Не правда ли, очень интересно, что Тутанхамон генетический европеец – удивляется Роман Шольц, директор центра iGENEA» (тот же сайт RusNews). Вот уж к кому не стоит обращаться за персональными интерпретациями своих ДНК данных (коммерческая компания iGENEA тестирует гаплотипы-гаплогруппы). Назвать Тутанхамона «генетическим европейцем» – это все равно, что назвать древних англичан «генетическими американцами», или «генетическими австралийцами».
Вот такая история про гаплотип Тутанхамона. Вероятность того, что он был правильно определен и описан (с экрана телевизора), есть немалая, но это нуждается в дополнительном подтверждении. Статья про египетские мумии к этому отношения не имеет.
Но что же еще та статья в Nature Communications показала, помимо определения трех гаплотипов, два из которых оказались недотипированы? В общем, мало что, во всяком случае для ДНК-генеалогии. Вот, например, показала, что если собрать все мумии, которые относились к до-птолемейскому периоду (XVI-IV вв. до н.э., то есть примерно 3600-2400 лет назад), птолемейскому (IV-I вв до н.э., то есть 2400-2100 лет назад) и римскому периоду (2050-1620 лет назад), и показать их распределение по митохондриальным ДНК (мтДНК), то получится следующая картина (две колонки справа – современные египтяне и эфиопы):
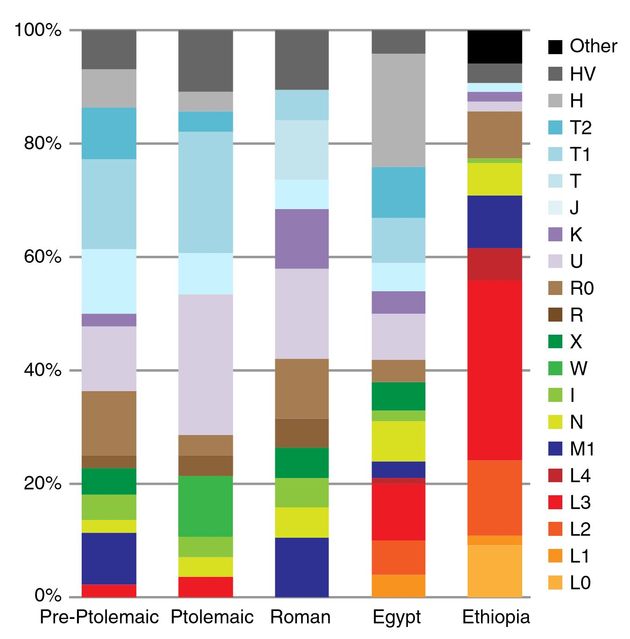
Информативно? Да не слишком, мягко говоря. Поэтому авторы статьи эту картинку даже не анализировали, привели просто так, типа «что вижу, то и пою». Да и в самом деле, что мы там видим? Что эфиопы от египтян по мтДНК далеки, как от древних, так и современных, а почему они должны быть близки? Эфиопов взяли только потому, что по ним есть данные, Эфиопия с Египтом даже не граничит (граничит с Эритреей, Джибути, Сомали, Кенией, Суданом и Южным Суданом). Да и какой смысл сравнивать древних египтян с современными эфиопами? Если авторы хотели показать, что ничего близкого нет, то в этом преуспели. Но является ли это решением исторической загадки? Вряд ли.
Еще нашли, что у современных египтян больше (по факту) «негритянской» гаплогруппы L (ее у эфиопов больше 60%), чем ее было у древних египтян. Иначе говоря, «понаехали». Является ли это исторической загадкой? Тоже вряд ли. Еще нашли (но в тексте не отметили, видимо, по причинам политкорректности), что в римском периоде негритянских (гаплогруппы L) мумий не обнаружено. У древнейших – нашли, у птолемейских – нашли, а вот при римлянах, из 19 мумий – никого. Вот такая незадача. А вот у современных египтян – не менее 20%. Историческая загадка?
Потом авторы статьи мтДНК вообще отложили, видимо, понимая, что выводы никакие, и переключились на «широкогеномные» данные. Они не так наглядны, как гаплогруппы, которые сразу видны. И что же они нашли? Что современные египтяне имеют на 8% больше африканской, суб-сахарской «предковости», чем древние египтяне всех трех периодов – древнейшего, птолемейского и римского. Ясно, что усредняли поперек всех, поскольку в римском периоде такой «предковости» вообще не было, но это обычное дело для попгенетиков. Погрешности для тех 8%, конечно, тоже не привели, да и кого это беспокоит, что там окажется 8±8%, не так ли? Что у современных больше – это и так видно, по мтДНК. Это и вынесли в заглавие статьи как самое большое достижение исследования.
Есть, правда, достижения меньшего масштаба, как следует из застенчивых описаний авторов, да и в заглавие не вынесли. Поскольку «широкогеномные» попгенетики оперируют размытыми понятиями «похожести», то и описания застенчивые. Вот, например, как геномы древних египтян, опять, конечно, усредненные поперек всех 90 образцов с разными гаплогруппами, соотносятся с другими древними популяциями, тоже, само собой, усредненными.
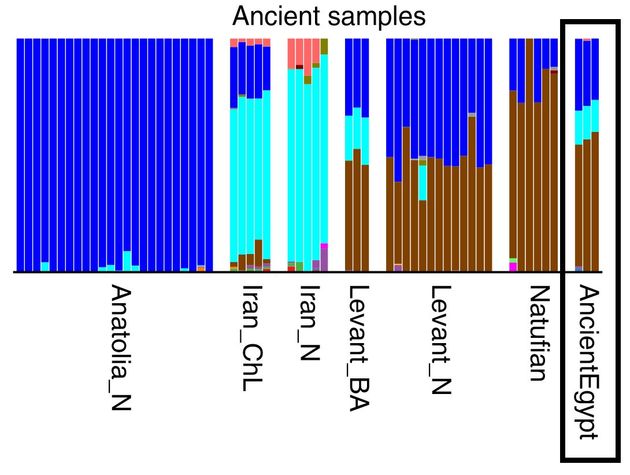
Видим, что в совокупных древних египтянах есть примерно треть от совокупных анатолийцев неолита, чуток от усредненных иранцев неолита и халколита, и больше половины то ли от натуфтйцев, то ли от неолитических левантийцев. Решение исторической загадки? Определили бы Y-хромосомные гаплогруппы – действительно решили бы исторические загадки. Но проблема в том, что «широкогеномных» попгенетиков исторические загадки не интересуют, их интересует просто описание того, что они увидели, а точнее – что посчитал компьютер. На самом деле уже три найденные гаплогруппы, даже в формате J, J и E1b, уже ясно показывают, что это потомки мигрантов с Ближнего Востока, для этого и полный геном не нужен, тем более, что он больше того все равно не показал, там данные конкретные, а здесь – вязкая «похожесть».
Хорошо, посмотрим, что дал усредненный геном древних египтян по сравнению с геномами современников Ближнего Востока и Северной Африки. Почему-то авторов статьи особенно интересовали геномы современных евреев. Хотели узнать, выходили ли древние евреи из Египта? В статье об этом ничего нет. Но геномы евреев есть – иракских, йеменских, ливийских, марокканских, тунисских. Африканских примесей (красный цвет – йоруба, негроиды западной Африки) ни у кого из них нет, с негроидами не смешивались, как и следовало ожидать. В статье этого тоже нет. Правда, и бедуины и друзы с теми негроидами не смешивались, хотя где Аравия (Иордания, Израиль, Ливан) и где западная Африка…
А так – все 18 усредненных геномов похожи (йоруба – 19-й), что так любят «широкогеномные», похожи что древние египтяне, что современные жители Ближнего Востока. Хотя это и из гаплогрупп Y-хромосомы очевидно. Таким образом, опять пришли к тому, что при наличии гаплогрупп Y-хромосомы «широкогеномные» данные ничего нового не дают, только последние значительно более неопределенные. А уж если гаплотипы добавить, то сравнения вообще никакого нет, это как «широкогеномный» плотник супротив столяра ©. Вот, собственно, и все основное содержание статьи в Nature Communications.
Продолжение следует…
Анатолий Алексеевич Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















