
В заглавие этой статьи можно было бы поставить «по молекулярной археологии человечества», поскольку археология, как известно, это наука, изучающая прошлое человечества по вещественным источникам. Поскольку «вещественные источники» в рассматриваемых в настоящей статье материалах – это ископаемые ДНК человека, то предмет исследования – источники молекулярной природы. Более того, мы выйдем за пределы археологии при обсуждении материала, потому речь пойдет о молекулярной истории, или, по-другому, о ДНК-генеалогии человечества.

В наших публикациях мы много раз говорили о том, что наступает время, когда историки поймут, что без данных ДНК-генеалогии их наука прогрессивно отстает от того, какой могла бы быть. К сожалению, среди современных историков только единицы поняли эту простую мысль. Подавляющее большинство хранят глухое молчание, а некоторые, напротив, заняли агрессивную позицию, «нападая» на ДНК-генеалогию и при этом показывая полную безграмотность, непонимание базовых основ новой науки. Можно было бы перечислить наиболее одиозных, но зачем? Им уже досталось на страницах «Переформата», в книге «Кому мешает ДНК-генеалогия» (2016, 845 стр.).
Но все-таки приведу недавний пример, когда некто И. Коломийцев, который называет себя историком (хотя другие историки это оспаривают, одна из цитат – «хоть и учился на истфаке Кубанского университета, но профессионально работал журналистом и занимался общественной деятельностью и политикой»), упоминает, что ДНК-генеалогия в моем лице якобы описывает «вечно победоносных славяно-ариев». Здесь Коломийцев исказил вдвойне – во-первых, никогда, ни в одной моей статье или книге нет оборота «славяно-арии», и я всегда подчеркиваю, что славяне и арии принадлежат разным эпохам; во-вторых, нигде и никогда у меня не было про «вечно победоносных» ни славян, ни ариев. У меня просто нет об этом сведений, напротив, я многократно писал, что арии были изгнаны из Европы и перешли на Русскую равнину в начале III тыс до н.э., около 5 тысяч лет назад. Балтийские славяне были, к сожалению, в значительной части онемечены еще в средние века, что «победоносностью» никак не назвать. Возможно, что Коломийцев не читал моих статей и книг, и пишет свою ерунду просто понаслышке, или вследствие агрессивной глупости. Но тогда особенно занятно, что Коломийцев жалуется – «пока всё, что я получаю — нападки и откровенные угрозы… под видом критики от «серьёзных учёных», не соизволивших даже прочесть мои произведения». Выходит, он не только лжец, но и лицемер. А поскольку, по его словам, его критикуют и «со стороны так называемых “патриотов”», то он, похоже, еще и русофоб. Язвительный оборот про «вечно победоносных» в отношении славян (или ариев) выдает его с головой.
Вот такие, с позволения сказать, «историки». Из всех исследований по истории ариев и истории славян, как и истории других этносов и народов, в том числе с привлечением ископаемых ДНК (гаплотипы и гаплогруппы Y-хромосомы) и рассмотрения соответствующих данных археологии и генезиса археологических культур в свете данных ДНК-генеалогии, их внимание привлекли только «славяно-арии», которых они сами же и выдумали. Страшно далеки они от науки.
На самом же деле место и роль ДНК-генеалогии в пересмотре и дополнении многих выводов истории и археологии, в ряде случаев и языкознания, описаны в десятках научно-популярных и научных статей в период 2009-2017 гг., а также в нескольких книгах, последняя – «История ариев и эрбинов» (2017), в которой подробно описаны и проанализированы результаты исследований десятков ископаемых ДНК, и обнаружено немало нестыковок и явных ошибок в академических интерпретациях археологических данных. Как я не раз подчеркивал, ДНК-генеалогия не ставит своей целью занять место археологии, истории, антропологии, языкознания, или потеснить их. Ничего подобного. У ДНК-генеалогии своя методология, свой характер описания результатов и выводов, свой расчетный аппарат, зачастую своя терминология, особенно когда принятая терминология является фактически устаревшей, ограниченной, или в ней просто нет терминов для явлений и событий, которые открывает ДНК-генеалогия.
Но поскольку подавляющее большинство археологов и историков пока бездействуют, тем самым откровенно устраняясь от рассмотрения вклада ДНК-генеалогии в свои дисциплины, и тем самым фактически предавая свои дисциплины, ДНК-генеалогия вынуждена заполнять вакуум. Это происходит с надеждой, что когда-нибудь, желательно в скором будущем, археологи и историки, а также лингвисты поймут, что время устаревших представлений уходит, и пора браться за работу, наверстывать упущенное. Появятся новые члены Национальных Академий наук за работы с привлечением ДНК-генеалогии. Это неизбежно, и первые выборы уже произошли – автор этой статьи избран в Национальную Академии наук Грузии (НАНГ) именно после доклада на Президиуме НАНГ по ДНК-генеалогии народов Кавказа. Представление произошло сразу двумя Президентами Академии – Т.В. Гамкрелидзе (недавно оставившим пост) и Г.И. Квеситадзе (недавно избранным Президентом), и затем (через два месяца) единогласно одобрено Общим собранием Академии. И хотя избранный академик числится по отделению биохимии, просто потому, что отделения ДНК-генеалогии в Академии пока не существует, да и потому, что по признанию Академии он внес крупный вклад в биохимию, в частности, в медицинскую биохимию (о чем и была большая инаугурационная статья в Бюллетене НАНГ), все в НАНГ понимали, что главным на выборах был вклад в ДНК-генеалогию. Пишу об этом потому, чтобы дать пример молодым (и не обязательно молодым) исследователям. С почином, так сказать.
На этой оптимистической ноте возвращаемся к основной теме статьи. В первые два месяца 2017 года прошел буквальный шквал статей по ископаемым ДНК, опубликованы десятки и сотни новых данных по гаплогруппам-субкладам в различных регионах Европы. Это позволяет получить значительно более продвинутую картину о миграциях древних народов и их потомков, о формировании новых популяций, часть из которых продолжают жить в виде этносов и в наше время, о том, кто в родовом отношении были обитателями ряда древних археологических культур, порой о взаимосвязи между этими культурами. Впрочем, для этих публикаций характерны и недостатки – авторы статей, как правило, генетики, не владеют ДНК-генеалогией, и потому либо вообще не публикуют гаплогруппы-субклады (про гаплотипы и говорить не приходится, их рассмотрение генетиками и попгенетиками практически никогда не проводится, они просто не знают, как это делать, ограничиваясь в лучшем случае расчетами их «разнообразия», никому и никогда не нужное), или выводят их в «Приложения», просто для упоминания. По той же причине авторы статей не владеют достаточно полной картиной современных представлений ДНК-генеалогии о миграциях носителей гаплогрупп по Евразии, у них отсутствует целостность представлений, и они в лучшем случае просто приводят сведения о гаплогруппах-субкладах, найденных в той или иной археологической культуры. Это на самом деле наиболее ценные у них сведения, другого от них и не надо. Но в худшем случае гаплогруппы-субклады не приводят, ограничиваясь некими диаграммами «метода принципиальных компонент», которые практически всегда приводят авторов либо к вязким неопределенностям, либо к принципиально неверным представлениям, противоречащих другим, несравненно более надежным данным и выводам, о которых авторы, судя по описанию, не имеют представления.
Поэтому прежде чем начинать приводить данные по гаплогруппам-субкладам (там, где гаплотипы не определялись, мы будем сообщать), дадим краткую сводку о том, что уже было известно (и что неизвестно) к настоящему времени. Тогда будет понятнее значимость новых находок ископаемых ДНК – что они подтвердили из того, что уже было известно или предполагалось ранее («на кончике пера»), что явно противоречит тому, что было найдено или предположено ранее, и что из найденного является принципиальным и долгожданным продвижением науки вперед. Иначе говоря, как изменилась картина наших представлений о древнем мире.
В этом разделе мы приведем основные результаты наших исследований, опубликованные в период с 2008 до 2017 гг., иногда в сопровождении результатов других исследователей (как правило, с ископаемыми ДНК). Общий предок современных носителей неафриканских гаплогрупп жил 64±6 тысяч лет назад, после прохождения бутылочного горлышка человечеством (в том числе, видимо, и неафриканским) в результате катастрофы планетарного масштаба. Это – наиболее вероятный сценарий, хотя другие пока не исключены, но они пока не предлагались с достаточными основаниями. Этот предок имел бета-гаплогруппу, или гаплогруппу ВТ, от которой образовались все гаплогруппы от В до Т в период времени от указанного до примерно 43 тысячи лет назад (гаплогруппа Т) и 37 тысяч лет назад (гаплогруппы N и О). Самые представленные в численном отношении в Европе гаплогруппы R1a и R1b образовались примерно 24300 и 21300 лет назад, соответственно, при определенных погрешностях расчетов (в линии гаплогруппы R1a среди ста человек имеется от 156 до 187 снипов на фрагмент нуклеотида размером 8.57 млн нуклеотидов, и в линии гаплогруппы R1b имеется от 129 до 165 снипов, что при 144 годах на снип дает приведенные датировки). Это подробно объяснено в Лекции 6 (в курсе из 18 видео-лекций). Коллектив YFull округляет различия и приводит датировки 22800±2300 лет назад для обеих гаплогрупп, R1a и R1b.
Гаплогруппы R1a и R1b образовались, видимо, в Южной Сибири, хотя прямых данных к тому нет, хотя их предковая гаплогруппа R найдена в ископаемых костных останках в Южной Сибири (рядом с Байкалом) с датировкой 24 тысячи лет назад, и там же, на Ангаре, найдена ископаемая гаплогруппа R1a с датировкой примерно 8500 лет назад. Гаплогруппа R1a с относительно близкой датировкой 7265±250 лет назад была найдена в Карелии, на Южном Оленьем острове. Все эти датировки не противоречат друг другу, как и тому, что носители гаплогрупп R1a и R1b пришли длинными миграционными путями из Южной Сибири в Европу. Детали миграционных путей обеих гаплогрупп по Евразии пока остаются неизвестными, но в общих чертах их можно изложить следующим образом.
По современным представлениям ДНК-генеалогии носители гаплогруппы R1a пришли в Европу по «южной дуге», через Иранское плато и Анатолию, прибыли на Балканы примерно 9-8 тысяч лет назад, расселились по Европе, и около 5000 лет назад, после (или в процесе) разделения на две основные ветви арийского субклада R1a-Z645, который образовался примерно 5500 лет назад, прибыли на восточную сторону Карпат и расселились по Восточно-Европейской равнине. С этой датировкой согласуется время расхождения индоевропейских языков на ветви. Эти ветви, в первую очередь, могли представлять северно-арийская (R1a-Z280) и южно-арийская (R1a-Z93) популяции. Первые остались на Русской равнине, вторые разошлись примерно 4500 лет назад по разным направлениям – южное, через Кавказ, которое примерно 4000 лет назад образовало митаннийских ариев (часть современной территории Сирии), хеттов (прилегающая и отчасти пересекающаяся с первой часть Анатолии); юго-восточное, в направлении Средней Азии и гор Таджикистана, с последующим переходом (примерно 3500 лет назад) на Иранское плато; восточное, в направлении Южного Урала и БМАК (Бактрийско-Маргианский археологический комплекс, 4200-3800 лет назад), и далее с переходом в Индостан примерно 3500 лет назад. Датировки ископаемых носителей гаплогруппы R1a в синташтинской культуре составляют 4030±115 и 4190±130 лет назад, датировка древнего поселения, южноуральского Аркаима (название современное), составляет 3800-3600 лет назад, по данным археологов.
Все эти датировки в целом согласуются друг с другом, хотя детальный маршруг перехода ариев в Индию остается неизвестным (возможно, через территорию современного Афганистана, возможно, через БМАК, возможно, другими путями), и археология перехода ариев в Индию пока отсутствует. Все ископаемые ДНК на этом пути (включая срубную, потаповскую, андроновскую археологические культуры), характеризуются гаплогруппой R1a-Z93 и ее нижестоящими субкладами Z94, Z2123, Z2124, или субкладом R1a-Z645, видимо, недотипированным. Практически все носители гаплогруппы R1a в современной Индии имеют субклады Z2123 и L657. Если маршрут первого уже хорошо известен, он пришел с северо-запада и севера, это субклад срубной культуры и затем синташтинской, то ископаемые L657 пока не найдены. Не исключено, что субклад R1a-L657 прибыл из стран Ближнего Востока, где среди арабов в настоящее время он широко распространен. Правда, небольшие количества L657 найдены в Казахстане и Иране, но туда они могли попасть обратным путем, из Индии или с Ближнего Востока.
Наконец, еще одна протяженная миграция ариев прошла в Зауралье – до Алтая (обычно субклад R1a-Z93 и нижестоящие Z2124, как в карасукской культуре, и R1a-Z645, видимо, недотипированные) и до Китая. На Алтае – пазырыкская культура (современные жители тех мест имеют в основном гаплогруппу R1a-Z93), там же ареал возникновения культур скифского круга. Оттуда скифы, носители (в частности) R1a-Z93, кочевали до Черноморского побережья и обратно, по степным лугам и лесостепным полосам Евразии. Скифы, конечно, имели не только гаплогруппу R1a, у них были разнообразные гаплогруппы, такие, как R1b, J2, G2a, Q, и говорили они, определенно, не только на языках индоевропейской группы, но и на тюркских языках. От них как ИЕ языки, так и тюркские языки, пришли на Кавказ.
Недавно были изучены два образца ископаемых ДНК из хазарских захоронений, датированных концом VII – началом VIII вв., и концом VIII – началом IX вв., оба имели гаплогруппу R1a-Z93 (Klyosov, Faleeva, Adv. Anthropol., 2017).
В наших статьях неоднократно предполагалось, что носители гаплогруппы R1a с древнейших времен, а именно со времен выхода на Русскую равнину, около 5000 лет назад, обживали побережье Балтийского моря. Там они намного позже встретились с носителями гаплогруппы N1a1 (старое название N1c1), которые вышли на Русскую равнину со стороны Урала 3000-3200 лет назад, и вряд ли ранее 3500 лет назад, и прибыли на берега Южной Балтики в середине I тыс. до н.э., то есть примерно 2500 лет назад, и на территорию современной Финляндии 2000-1500 лет назад. Никакого «финно-угорского субстрата» на Русской равнине ДНК-генеалогия не находит, равно и никакой «ассимиляции» «местных финно-угров прибывшими славянами». Это – пунктик у Балановского, о чем он не устает говорить, как и опять в недавней радиопередаче. Ну не нравятся ему славяне, вот и придумывает для них всяческие «политически некорректные» действия, к восторгу либеральных кругов, но не давая никаких научных обоснований.
Именно потому, что носители гаплогруппы N1a1 прибыли на Русскую равнину (включая ее север) довольно поздно, намного позже, чем носители R1a, то для меня всегда было удивительно, как попгенетики вместе с немногими историками отдают культуру ямочно-гребенчатой керамики гаплогруппе N1a1. Данных на этот счет вообще не было, поэтому не стоило было вообще на этот счет высказываться, тем более «по понятиям». Но это уже было в Википедии, опять, конечно, с подачи попгенетиков, поэтому не менять же тем же попгенетикам. На самом деле это до последнего времени оставалось неизвестным, а как там было – будет описано и систематизировано в настоящей статье.
Гаплогруппа R1b пришла в Европу по «северной дуге», со стороны Южной Сибири (хотя прямых доказательств и деталей миграции опять нет), через Южный Урал, Северный Казахстан, культуры Тургайского прогиба, образуя по пути многочисленные археологические культуры, в том числе терсекская, суртандинская, маханджарская, ботайская, хвалынская, самарская, средневолжская, ямная, ранняя катакомбная и другие. Прямых доказательств, что это были культуры эрбинов, носителей R1b, не было, но были многочисленные косвенные доказательства (как, например, положения костяка на спине), подробно, со многими примерами изложенное на Переформате в статье (в двух частях) «Как Западную Европу заселили новые европейцы» еще три года назад. Это всё, с перечислением культур, включая хвалынскую и ямную, было изложено в статье «Древняя история эрбинов» в журнале Advances in Anthropology еще в 2012 году, пять лет назад, и в недавней книге «История ариев и эрбинов» (2017). Многие отнеслись к этому с недоверием, попгенетики, ссылаясь на историков, утверждали, что это невозможно, что насельники ямной культуры были индоевропейцами, следуя теориям Гимбутас, а значит, у них должна была быть гаплогруппа R1a, и так далее. Все возражения рассыпались в 2015 году, когда было опубликовано (Haak, Nature, 2015; Allentoft, Nature, 2015), что одиннадцать ископаемых ДНК ямной культуры имели гаплогруппу R1b (плюс одна забредшая гаплогруппа I2a в Калмыкии, c датировкой 4510±350 лет назад). В хвалынской культуре у трех образцов оказались три разных гаплогруппы – R1b, R1a и Q1a, все с археологической датировкой 6615±600 лет назад, что для арийской R1a совершенно нехарактерно, датировка слишком древняя. Иначе говоря, этот образец не из тех R1a, что пришли в те края через две тысячи лет из Европы, и не удивительно, что он был погребен по обычаям эрбинов, на спине.
Оказалось, что доминирующим субкладом в ямной культуре был R1b-Z2103, археологическая датировка 5300-4500 лет назад, и его носители сейчас во множестве живут на Кавказе, это там основной субклад гаплогруппы R1b. Через Кавказ его носители прошли в Месопотамию, и в обход Черного моря прошли на Балканы, разойдясь по юго-восточной Европе. В центральной и западной Европе его почти нет, там доминируют носители нижестоящих субкладов гаплогруппы R1b-L11, потомков культуры колоколовидных кубков.
Гаплогруппы G2a и I2 доминировали в Старой Европе, то есть до середины III тыс. до н.э. (примерно 4500 лет назад), затем практически исчезли, как, впрочем, и европейские носители гаплогрупп R1a и E1b. Все они прошли бутылочное горлышко популяции примерно 4500 лет назад, и возродились из немногих, видимо, выживших носителей этих гаплогрупп, или небольшое время спустя, но на периферии Европы, G2a – в Малой Азии и на Кавказе; I2 – на Британских островах и намного позже на Дунае-Карпатах, R1a – на Среднерусской возвышенности, E1b – на Балканах, ареал возрождения гаплогруппы I1 неясен, их гаплотипы практически одинаковы по всей Европе, но наибольшее их количество сейчас в Скандинавии – неизвестно, они там возродились, или туда переместились примерно 3700 лет назад из какого-то другого места.
Вот так представляется картина перемещений, миграций племен и народов по современным представлениям, без учета новых публикаций первых двух месяцев 2017 года. Рассмотрим, как новые данные изменили эту картину.
археологические культуры ямочно-гребенчатой керамики (6200-4000 лет назад)
и шнуровой керамики (5200-4300 лет назад)
Конечно, «ископаемые гаплогруппы» в названии данного раздела – это «скоропись», более правильно было бы написать «ископаемые костные останки, из которых была извлечена ДНК, в которой были выявлены специфические мутации гаплогруппы R1a». Но каждый раз повторять это неудобно, так что давайте примиримся со скорописью. В научных описаниях часто применяют подобную скоропись и даже многобуквенные сокращения.
Поясним, что культура ямочно-гребенчатой керамики (КЯГК, иногда называют «гребенчатой керамики») – это археологическая культура (точнее – «культурно-историческая общность») каменного века на северо-востоке Европы, ее обычно датируют временами 6200-4000 лет назад; в статье, которую сейчас рассмотрим, для нее приводят датировки 5900-3800 лет назад, видимо, потому что именно таковы археологические датировки костных остатков, в которых анализировали ДНК. На карте, приведенной в Википедии, регион этой культуры расположен в верхней правой части. Видно, что туда попадает часть территории современной России, Белоруссии, Прибалтики.

Не очень понятно, почему, но популяционные генетики настойчиво приписывали КЯГК гаплогруппу N, называли культуру «финно-угорской», и цитировали немногих антропологов, которые утверждали, что насельники этой культуры относились к уральской расе, и в их ископаемых скелетах «обнаруживали монголоидные элементы». Правда, в Википедии после этих описаний есть примечание «неавторитетный источник», и стоит требование предоставить дополнительную информацию, которая так и не была представлена. Поэтому все, видимо, попгенетики ожидали, что ископаемая ДНК из КЯГК покажет гаплогруппу N (с соответствующими подгруппами), и они, попгенетики, опять будут повторять про «финно-угорский субстрат» по всей Русской равнине, и про то, что пришли славяне и всех ассимилировали. В том числе и эстонцев, и литовцев с латышами, у которых всех примерно по половине мужского населения имеют гаплогруппы R1a и N1a1, но R1a – это, само собой, гаплогруппа якобы оккупантов.
Разбор статьи. В марте 2017 года вышла статья Saag c коллективом в 14 авторов, из которых более половины – сотрудники научных центров Эстонии, хотя выделение и анализ ископаемых ДНК проводили в Центре геогенетики в Дании под руководством уже знакомого нам Eske Willerslew. Статья опубликована как препринт в издании bioRxiv, но нет больших сомнений, что она вскоре появится в Nature. Название статьи – «Экстенсивное фермерство в Эстонии началось путем миграций из Степи, несбалансированных в половом отношении». Название, надо сказать, весьма бестолковое, особенно учитывая то, на чем было основано это «несбалансированное». А основано это было на том, что достаточно произвольный анализ фрагментов ДНК, так называемый «outgroup f3 анализ», дал ряд выпадающих точек, что было интерпретировано авторами так, что распространение культуры шнуровой керамики (КШК) в Эстонию указывало на мужских предков из Степи, а женских предков – на ранних европейских фермеров (или фермерш). Вот и всё, авторы решили, что мужчины «несли гены» из степи, а женщины – из Европы, и авторы внесли это в название статьи, так высоко оценили эту «находку». Видимо, посчитали самым важным во всей работе. Хотя можно было бы подумать, что при датировках КШК в Эстонии от 4800 лет назад вкупе с гаплогруппой R1a никакой «степи» там быть просто не могло. КШК распространялась из Европы начиная примерно с 5200-5000 лет назад, и в Эстонии была еще на ранней стадии. В степи они ушли, опять же из Европы, уже заметно позже. Так что с гаплогруппой R1a «из степи» 4800 лет назад, да и позже, авторы определенно ошиблись. Это не единственная ошибка в статье, там их много, о чем упомянем ниже.
На карте, взятой из статьи, указаны захоронения, из которых извлекли ископаемые ДНК. Слева цветными контурами показаны части ареалов культуры шнуровой керамики (красным цветом), причем двойной красной окантовкой показан «эпицентр» захоронений, отнесенных к данной культуре; синим цветом – культуры ямочно-гребенчатой керамики, и серым, в самой нижней части карты, нарвской культуры (захоронения с датировкой 6400-6200 лет назад), в которой, впрочем, из найденных археологами 26 скелетов был выбран для исследований только один, скелет девочки, и в нем нашли мтДНК U5a2d. Это мало о чем говорит, поскольку U5 – это типичная митохондриальная гаплогруппа Евразии древнейших времен. Справа на карте отмечены места захоронений (в том числе и тех, которые в статье не изучали), теми же цветами: темно-серым – нарвской культуры, темно-синим – культуры ямочно-гребенчатой керамики, темно-красным – культуры шнуровой керамики. Карта в увеличенном формате открывается в новом окне по клику:
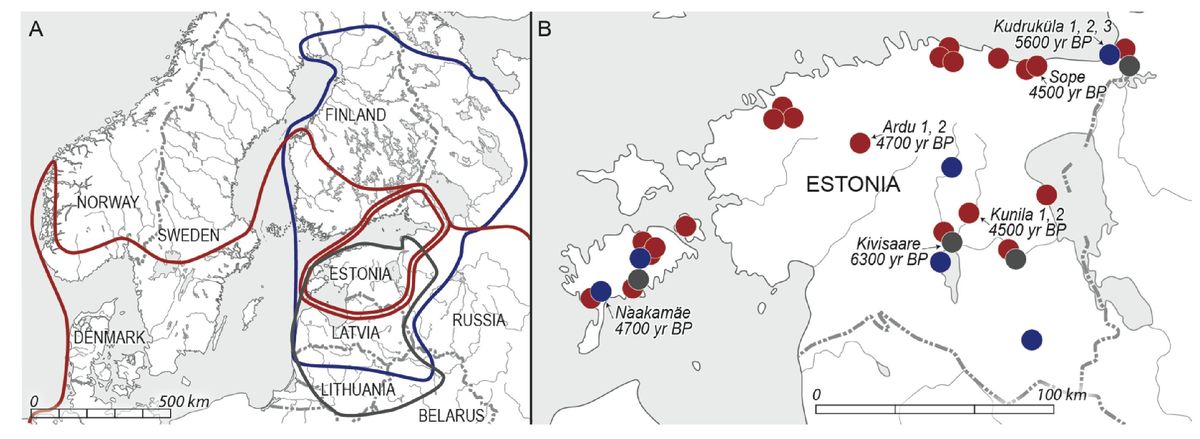
Мы кратко перечислим гаплогруппы с субкладами, которые нашли авторы. Остальное, что описывают авторы статьи, не стоит того, чтобы на этом останавливаться. Как обычно, это вязкие, неуверенные рефлексии, которые уже стали визитной карточкой не только самих авторов, но попгеномной тематики в целом. И понятно, почему вязкие и неуверенные. В отличие от четких и однозначных показателей ДНК-генеалогии – конкретной гаплогруппы, соответствующего субклада (хотя субклады часто бывают недотипированы), конкретного гаплотипа (хотя гаплотипы бывают относительно короткие или протяженные), в данной статье, как и других подобных, идет сплошной произвол как в представлении, так и рассмотрении данных. Вот – простой пример. На диаграмме приведены «результаты анализа ADMIXTURE при K14 для 225 древних образцов ДНК», где «ADMIXTURE» – это кодовое слово, означающее «примесность». Мы сейчас увидим, что оно дает. А К14 – это произвольно выбранная степень разложения геномов, усредненных или индивидуальных, на 14 компонентов. Такое число компонентов – потому что так решили. Могли бы выбрать любое число компонентов, скажем, 8, или 6, или 10, и в каждом случае диаграмма была бы другой. Выбранное число считается высокой степенью разложения.

Цвета на диаграмме – это «референсные» геномы, тоже усредненные, поперек всех гаплогрупп, поперек геномов мужчин и женщин, без разбора. Взяли древние геномы, назвали их, например, «степные EMBA» (это темно-красные на диаграмме), степные MLBA» (светло-красные), «степные EN» (голубые), «европейские LNBA» (розовые), «анатолийские/левантийские EF» (темно-зеленые), «европейские EF» (светло-зеленые), «Европейские HG» (голубые), «Степные EN» (тоже голубые), «кавказские/иранские HG» (серо-зеленые), «палеолитические HG» (темно-серые), усреднили в каждой категории, и нанесли на диаграмму ниже. EMBA – это ранний-средний бронзовый век, MLBA – средний-поздний бронзовый век, LNBA – поздний неолит – бронзовый век, EN – энеолит, EF – ранние фермеры, HG – охотники-собиратели. Усредняют, повторяю, без разбора, чохом. Например, в хвалынской культуре нашли три образца ДНК, один гаплогруппы R1b, другой – Q1a, третий – R1a. Все три усреднили, разложили по цветам, назвали Samara_Eneolithic_av, где av – это и есть «average», то есть усредненный), он оказался точно таким же, как для срубной культуры, андроновской, синташтинской, шнуровой керамики, то есть больше, чем наполовину синий, на треть зеленый, на одну десятую темно-красный, и по крохам светло-зеленый и еще пару совсем минорных цветов. Но проблема в том, что срубные, андроновские, синташтинские, шнуровики – все R1a, так они и были определены для использованных здесь ископаемых ДНК. А хвалынский – усреднен по трем разным гаплогруппам, и всё равно получилось то же самое. Ну и что это дает? Как что – по ним «происхождение» устанавливают. Сравнивают, и если похожи, значит, происхождение одинаковое. Представляете?
Короче, все они на 50-60% «европейские охотники-собиратели» (голубые), они же «степные энеолитические», на 20-30% «анатолийские-левантийские ранние фермеры», на крошку «степные ранний-средний бронзовый век», еще на крошку – «ранние (европейские) фермеры». Представляете? И ямная культура, и хвалынская, и европейская культура шнуровой керамики, и вообще практически все. И шнуровая керамика Германии, и колоколовидные кубки, и синташтинцы, и вообще все. И знаете, почему? А потому, что сказать ничего более определенного просто невозможно, методология такая. Вязкая, расплывчатая, уклончивая.
Посмотрим еще раз на диаграмму. Там все практически одинаковы – и геном афанасьевской культуры (опять «усредненный»), и полтавкинской (R1b), и потаповской (R1a), и ямной (R1b, как в Самаре, так и в Калмыкии), и андроновской (R1a), и синташтинской (R1a), и срубной (R1a), и культуры колоколовидных кубков (R1b), и полтора десятка образцов культуры шнуровой керамики (все R1a), и эстонские образцы культуры шнуровой керамики (все четыре R1a), и один женский (Sope), в котором никаких мужских гаплогрупп нет и быть не могло – и опять идентичная диаграмма, как у всех прочих. Так что эта диаграмма показывает? Да, в общем, что-то показывает, она же объективная реальность, данная нам ощущением, зрительным во всяком случае. Только что показывает – так это откровенную профанацию. По сути – ничего не показывает, но обсуждается на многих страницах статьи, делаются выводы, хотя можно без затруднений делать и противоположные. Вот авторы и пишут (здесь умышленно дается буквальный перевод – оцените обычную для подобных статей вязкость теста):
«Контрастирующая картина частоты аллелей аутосом по сравнению с Х-хромосомными при аутгруппном анализе f3 показывает, что Х-хромосомы культуры шнуровой керамики намного более подобны популяции ранних фермеров как в Анатолии/Леванте, так и в Европе. Однако на аутосомах мы видим противоположное – большинство европейских охотников-собирателей /степных энеолитических популяций, степных EMBA и CHG/древних иранских популяций имеют тенденцию к более высоким величинам f3».
Кто что понял? И вот так – сплошная статья. Ладно, обсуждать это никакого смысла не имеет. Ни на один вопрос, который можно поставить, статья ответов не дает. Кстати, статья, приведя в таблице гаплогруппы шести образцов (один из культуры ямочно-гребенчатой керамики, и пять из культуры шнуровой керамики), их практически не обсуждает, и тут же переключается на бессмысленные по сути обсуждения диаграммы, приведенной выше. В частности, опять о том, что «диффузия из степей [ссылка на М. Гимбутас, 1956 год], как недавно было поддержано изучением аутосомных ДНК [ссылка на «широкогеномные» статьи 2015 г, Haak и др. и Allentoft и др., Nature] показало ассоциацию с появлением культуры шнуровой керамики в Европе». Замечаете уклончивый стиль, типичный для попгенетиков? «Показало ассоциацию с появлением культуры…». Я уже не раз цитировал директора института археологии в США, которая назвала фантастической безграмотностью положение упомянутых «широкогеномных статей», авторы которых связали ямную культуру с появлением культуры шнуровой керамики в Европе. Это было в 2015 году, и исходная статья, и критика (сначала на Переформате, затем американским археологом) давно прошла, сейчас уже 2017 год, но авторы, видимо, необучаемы.
Так что там с гаплогруппами Y-хромосомы? В нарвской культуре не определяли. В ямочно-гребенчатой культуре определяли в одном образце (с датировкой 5900-3800 лет назад), гаплогруппа R1a-M459-YP1272, в пяти образцах культуры шнуровой керамики (датировки 4800-4500-4300 лет назад, определены радиоуглеродным методом) – все R1a-Z645, но любители из сети разобрались с их геномами в базе данных, и сообщили, что в двух случаях из пяти Z645 – это недотипировано, должно быть Z283 и Z282, то есть дополнительные нисходящие снипы. В захоронении Sope (северо-восток Эстонии, см. карту выше) изучили два скелетных материала, обе оказались женщинами, гаплогруппы мтДНК H5a и H5a1, датировки 4575-4350 лет назад. Но авторы пишут, что там найдены захоронения 10 человек, все лежат в скорченном положении на боку, что, как пишут авторы, характерно для Эстонии и для всего восточно-балтийского региона. Но это (о чем авторы не пишут) – характерное положение костяка для носителей гаплогруппы R1a, мужчины на правом боку, голова в направлении на запад, женщины на левом боку, голова на восток, все лицом на юг. Таким образом, есть основания полагать, что в культуре шнуровой керамики восточно-балтийского региона конца – середины III тыс. до н.э. среди мужчин преобладали (или были практически исключительно) носители гаплогруппы R1a. Вот, собственно, и все, что в статье стоит читать. И статья сразу становится значимой.
Она значима, потому что ее экспериментальные данные отвечает на несколько важных вопросов. Они (данные) опровергают «положение», которого держались попгенетики много лет, что ямочно-гребенчатая культура была исключительно культурой носителей гаплогруппы N. Это было краеугольным камнем «положения» о «финно-угорском субстрате» на Русском Севере и на Русской равнине, и о преобладающем населении там гаплогруппы N. Ничего из этого не подтверждается, куда ни посмотри. Например, в межовской культуре позднего бронзового века, на севере Челябинской области, которую историки ранее дружно относили к «уграм», вместо гаплогруппы N найдены ископаемые гаплогруппы R1a-M417 и R1b (в Каповой пещере).
Далее, они опровергают «положение», что насельниками Балтийского побережья с древнейших времен были «финно-угры», что попгенетики немедленно превратили в «положение» о гаплогруппе N в тех краях, и об «ассимиляции» их пришедшими славянами, или в более общем виде – пришедшими носителями гаплогруппы R1a. Оказалось – не так, а что носители R1a жили там с древнейших времен, во всяком случае с 5-6 тысяч лет назад. В соседней Карелии найдена гаплогруппа R1a-M459-YP1272 с датировкой 7265±250 лет назад, тот же субклад, что и в культуре ямочно-гребенчатой керамики в Эстонии. В Германии (в Эулау) в культуре шнуровой керамики еще в 2008 году была выявлена гаплогруппа R1a, с археологический датировкой 4600 лет назад. Всё это образует поле обитания носителей гаплогруппы R1a от южной Германии к северу и востоку, по Прибалтике, что и заложило равное количество гаплогруппы R1a с гаплогруппой N1a1 у современных литовцев, латышей, эстонцев, и далее мы видим переход к югу с включением фатьяновской археологической культуры (4300-3500 лет назад), древних русов, и южнее, вплоть до Черного моря, с обитающими там носителями гаплогруппы R1a-Z93 и определенно других субкладов. Этого, конечно, в рассматриваемой статье нет.
Чтобы не сложилось впечатление, что я предвзято отнесся к разбираемой здесь статье, процитирую целиком «Заключение» к статье. Пусть каждый делает выводы о его стиле и значимости, но помнит, на чем эта цитируемая «значимость» основана, и еще раз посмотрит на цветную диаграмму выше.
Наши результаты поддерживают гипотезу, что индивидуалы, ассоциируемые с охотниками-собирателями культуры гребенчатой керамики в Эстонии генетически наиболее подобны восточным охотникам-собирателям из Карелии, к востоку от Эстонии [Примечание – генетические характеристики образца из Карелии в статье вообще не упоминаются, и никаких сравнений с ними не проводилось – ААК]. Недавно опубликованные сравнения геномов мезолитической Латвии и охотников-собирателей культуры гребенчатой керамики поддерживают этот результат, в то время как мезолитические охотники-собиратели из Латвии и Литвы, видимо, генетически наиболее подобны западным охотникам-собирателям. Это свидетельствует о степени генетического вливания с востока при прибытии носителей культуры гребенчатой керамики. Далее, присутствие генетического компонента, ассоциированного с кавказскими охотниками-собирателями и позже с людьми, представляющими ямную культуру в индивидуалах восточных охотниках-собирателей и эстонских индивидуалах культуры гребенчатой керамики означает, что экспансия культуры шнуровой керамики не может рассматриваться как единственный способ распространения этого генетического компонента, по меньшей мере в Восточной Европе. Переход к интенсивному фермерству и животноводству в Эстонии, который произошел несколькими тысячелетиями позже перехода к фермерству во многих других частях Европы, проводился индивидуалами культуры шнуровой керамики и включал прибытие нового генетического материала. Эти люди привнесли очевидную степную предковость вместе с некоторым минорным анатолийским вкладом, наиболее вероятно абсорбированным через женские линии в ходе популяционных передвижений. Наши результаты подтверждают недавно сообщенное утверждение, что фермерство не прибыло в Балтийский регион как следствие ступенчатой миграции людей из Анатолии. Однако наши генетические данные, ассоциированные с археологическими сообщениями, демонстрируют, что переход к фермерству в Балтийском регионе не был также результатом только культурной трансмиссии, но вместо того был результатом миграции фермеров из Степи. Более того, поскольку генетическая вариация современных эстонцев не может быть полностью объяснена этими событиями, нужны дальнейшие исследования для выявления последующих демографических событий, которые принесли гаплогруппу N на берега Балтийского моря и изменили аутосомную вариацию людей, живущих в этом регионе.
Кто что понял? Можно было, конечно, подредактировать, но потерялся бы изыск авторского «Заключения». Может, кто что понял про фермеров из Степи? Или вообще, что авторы хотели выяснить, и чего достигли? И вот теперь сравните мой пересказ выше по гаплогруппам-субкладам Y-хромосомы, описанным в статье, и их значимости, в том числе и в более широком представлении, и совершенно мутное «заключение» авторов статьи. Вот так ДНК-генеалогия отличается от популяционной генетики вообще и «широкогеномной» попгенетики в частности.
культуре (7000-5000 лет назад), на территории современной Латвии
В феврале 2017 года в журнале Current Biology вышла статья под характерным названием – «Неолитический переход на Балтике не был вызван примесностью (admixture) с ранними европейскими фермерами» (Jones и др.). Характерное потому, что полностью противоречит выводам статьи, разобранной в предыдущем разделе. Это совершенно типично для «геномных» статей, которые трактуют полученные данные совершенно произвольно, «по вкусу» и «по понятиям» авторов, и скорее всего одного автора, который статью писал. Причины описаны выше, не будем их повторять. Данная статья строилась и обсуждалась по тем же принципам, что описаны выше. Отсюда и типичные «положения», цитирую: «анализ, проведенный с помощью (программы) ADMIXTURE предполагает близкое взаимоотношение между этими группами (WHG и EHG, то есть западно-европейские охотники-собиратели и восточно-европейские охотники-собиратели). Мы нашли, что хотя мезолитические образцы из Латвии имеют более близкую аффинность к WHG, чем к EHG, они не относятся полностью ни к одной из этих групп».
Информативно, не так ли? И вот так вся статья. Для сравнения, два из упомянутых мезолитических образцов, для которых были определены Y-хромосомные гаплогруппы, оказались R1b-P297, и это сразу выстраивает картину миграций. Об этом – ниже. Но «геномные попгенетики» гаплогруппы и субклады не анализируют, не говоря о гаплотипах. Потому и их многочисленные проблемы, недоговорки, заблуждения. Например, неолитический образец из Латвии (Y-гаплогруппа не указана, археологическая датировка 6000±200 лет назад) после проведения анализа с той же программой ADMIXTURE «показал», что «имеет несколько компонентов, которые были найдены среди американских индейцев, сибиряков и охотников-собирателей с Кавказа».
Информативно, не так ли? Непонятно, зачем все это, когда больше к этой «информации» авторы в статье не возвращаются. Что вижу, то и пою? ©
В статье опять повторяется, что гаплогруппа R1b « распространилась из России, с востока, в Европу после 5000 лет назад». Датировка правильная, но неправильное направление. Если бы она распространилась в Европу с востока, то как она бы попала на Пиренейский полуостров, откуда и двинулась в Европу как культура колоколовидных кубков? На самом деле этот вопрос давно проработан ДНК-генеалогией, и показано, что эрбины, носители гаплогруппы R1b, никак не могли двинуться в Европу напрямую с востока, они прошли туда, на Пиренеи, кружным путем. Эта ошибка геномных попгенетиков уже стала традиционной, повторяя «выводы» двух статей 2015 года, Haak и др., и Allentoft и др. А поскольку методология в статье, как и у других геномных попгенетиков, вся в прорехах, а местами и вовсе неверная, то ее нечего и обсуждать, это мы делали во многих других статьях. Перейдем к сути находок, а именно к гаплогруппам и субкладам. Гаплотипов в статье, конечно, нет.
ДНК восьми образцов были протестированы на мтДНК, и только два из них протестировали на Y-хромосомные гаплогруппы, оба из нарвской культуры, археологическая датировка 7690±100 и 7030±225 лет назад. Оба образца имели гаплогруппу R1b, и, как собщили авторы статьи, обе имели «как наиболее вероятный» субклад R1b1b, далее добавив, что эта гаплогруппа наиболее распространенная среди современных западных европейцев. Как следствие, здесь же было сообщена «интерпретация», что «присутствие этой гаплогруппы в мезолитической Латвии указывает на ее западное происхождение». Здесь – целый ряд принципиальных ошибок. Во-первых, популяционные генетики никак не поймут, что уже давно, лет десять, нельзя ограничиваться строчными наименованиями субкладов (как R1b1b), надо непременно указывать индекс снипа, иначе возникают множественные недоразумения и делаются неверные выводы. В данном случае – какого года номенклатуру использовали? В последней, 2017 года номенклатуре ISOGG указанный субклад числится как R1b1b-PH155, до этого шесть лет (с 2011 до 2016) это был R1b1b-M335, и в 2010 году это был R1b1b-P297. В статье никаких разъяснений нет. Тем временем энтузиасты-специалисты, известные в сети, провели свой генетический анализ по геномным базам данных, и установили, что этот образец относится к субкладу «pre-M73», то есть явно прибыл с востока, видимо, из южной Сибири, а другой образец, с датировкой 6060±140 лет назад, относится к субкладу R1b1a1a-P297 (см. диаграмму снипов ниже), то есть авторы статьи, возможно, просто использовали давно устаревшую номенклатуру (2010 года) с усеченным строчным наименованием субклада.
Но на этом недоразумения не останавливаются. Субклад Р297 является одним из старейших в системе субкладов гаплогруппы R1b, он образовался 16800±2600 лет назад (https://www.yfull.com/tree/R1b/), предположительно в южной Сибири (там же, на Байкале, нашли его родительскую гаплогруппу R с археологической датировкой 24 тысячи лет назад), и он никак не мог прибыть более семи тысяч лет назад с запада, из Европы, чтобы оказаться в нарвской культуре в Латвии. Известно только, что его дочерний субклад, R1b-P297 > M478 > Y14051, который образовался примерно 7200 лет назад, обнаружен у современного жителя Украины, и на нем этот субклад заканчивается, нижеследующих субкладов (и снипов) пока обнаружить не удалось. Вот так небрежность в отнесении субкладов, то есть использование устаревшей номенклатуры, ведет к принципиально ошибочным интерпретациям экспериментальных данных. Основные миграции носителей гаплогруппы R1b прибыли в Европу только около 5000 лет назад, и у них были уже намного более поздние субклады, которые в диаграмму ниже не внесены, они были бы далеко внизу. Конечно, возможны редкие приходы в Европу и более древних носителей гаплогруппы R1b, которые позже полностью вымерли вместе с потомками, но раз мы о них не знаем, то на них основывать ничего нельзя.
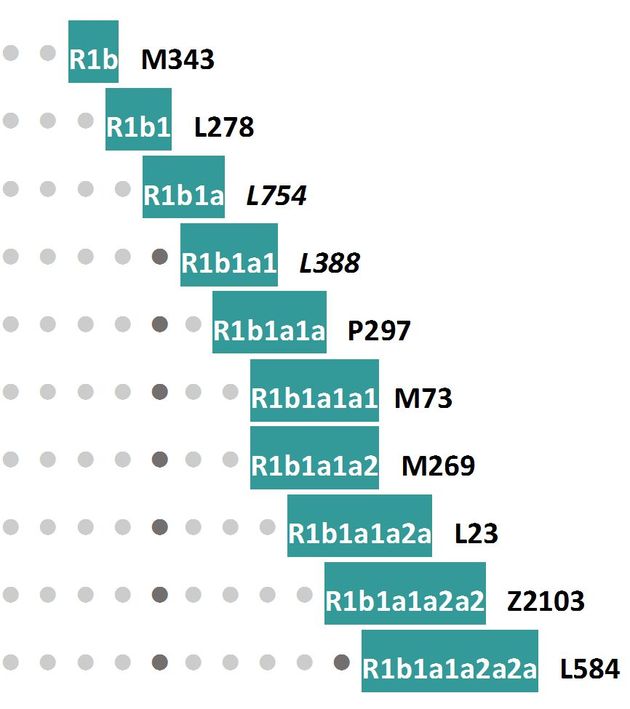
В любом случае, находка авторов статьи является очень важной (если не смотреть на неверные интерпретации авторов статьи), она впервые продвигает ареал миграций и обитания эрбинов от средней Волги (что показали изучения ископаемых ДНК ямной и хвалынской культур) до Балтики, причем для времен 7000-7700 лет назад, начала нарвской культуры. Эти времена пересекаются с таковыми для хвалынской культуры, в которой найдена та же гаплогруппа R1b, но более древнего субклада L278 (= M415), приведенного на диаграмме выше в самом начале, он образовался 20400±3000 лет назад. Иначе говоря, это родительская (по отношению к нарвским R1b) Y-хромосомная линия. Так что, похоже, что эрбины в период 7-8 тысяч лет назад передвигались по Русской равнине от Кавказа до Балтики, в те же времена там же (во всяком случае в Прибалтике) обитали и носители гаплогруппы R1a (культура ямочно-гребенчатой керамики, территория современной Эстонии и, возможно, всей юго-восточной части Прибалтики, включая Карелию, см. предыдущий раздел). Ямная культура, которая датируется историками/археологами как 5600/5200-4300 лет назад, представляет собой только «хвост» миграций эрбинов на Русской равнине, что давно показывали данные и публикации ДНК-генеалогии. В последующие времена эрбины или сами ушли на юг, на Кавказ и далее в Месопотамию, или их вытеснили арии, но итог мы знаем – на Русской равнине сейчас проживают примерно 50% носителей гаплогруппы R1a (до 67% в южной части Центральной России), и только 5% носителей гаплогруппы R1b. Возможно, потому историки пишут, что нарвская культура исчезла под натиском племён ямочно-гребенчатой керамики, и мы, уже зная, что первые – это носители R1b, вторые – носители R1a, понимаем, что это было началом крупных исторических процесов на Русской равнине.
В расматриваемой статье этих интерпретаций и выводов, конечно, нет. Можно упомянуть, что в рассматриваемой статье определены также мтДНК для восьми образцов ископаемой ДНК:
— два из них упомянутой нарвской культуры c Y-хромосомной гаплогруппой R1b (U5a2d и U2e1), 7690±100 и 7030±225 лет назад,
— один – для более древней культуры кунда (U5a1c), Латвия, 8300±100 лет назад,
— три – для неолитических культур Латвии (U4a1, U4, U5a1), соответственно 6100±100, 6000±200, и 4800±200 лет назад, последняя – культура шнуровой керамики, и
— два – для мезолитического и неолитического образцов ДНК днепровской Украины (U5b2 и U4), 10900±300 и 6380±90 лет, сответственно.
Для последних двух образцов энтузиасты из сети определили по геномным базам данных Y-хромосомные гаплогруппы-субклады, которые оказались соответственно I2a2a-M223 и R1a-M459. Первый – древний субклад, образовался примерно 17500 лет назад, был одним из наиболее распространенных в Европе до примерно 4500 лет назад, его носители были, видимо, в основном уничтожены при расселении по Европе эрбинов, культуры колоколовидных кубков, и общие предки современных M223 жили в основном на Британских островах, с датировкой примерно 5000 лет назад, то есть намного более недавних, чем был обнаружен на Украине. Но данные важны в том отношении, что показывают, что гаплогруппа I2 продвигалась со стороны центральной и западной Европы (например, Испания и Франция), а также Скандинавии, до территории современной приднепровской Украины. В таком случае не будет удивительным, если ископаемая гаплогруппа I2 будет найдена в культурах трипольской и сопряженных с ней. Второй оказалась гаплогруппа R1a-M459 (c датировкой 6380±90 лет назад), то есть уже неоднократно найденной доарийской R1a (если арийские отсчитывать со времени образования субклада R1a-Z645, примерно 5500 лет назад), которая обнаружена в хвалынской мезолитической культуре (R1a-M459), прибалтийском мезолите (R1a-M459-YP1272), в мезолите Карелии (R1a-M459-YP1272). Иначе говоря, складывается система, в которой доарийская R1a (образовалась ранее 6000 лет назад) была довольно широко представлена на Русской равнине, но в итоге не прошла бутылочное горлышко популяции – общий предок современных Y-хромосом на Русской равнине жил примерно 4900 лет назад, и относился, видимо, к субкладу R1a-Z280, на шесть ступеней ниже, чем М459 (R1a-M420 > M459 > M198 > M417 > Z645 > Z283 > Z282 > Z280).
Обращает внимание, что все восемь образцов имеют мтДНК гаплогруппу U, наиболее распространенную для мезолитической Европы. Большего, пожалуй, из этих гаплогрупп-субкладов не извлечь, как и из того, что для серии ископаемых ДНК в предыдущем разделе:
— образец ДНК нарвской культуры имел мтДНК U5a2d (6300±60 лет назад),
— четыре образца из культуры ямочно-гребенчатой керамики имели U5b1d1 (дважды), U4a и U2e1, с датировками 5600±200, 5600±200 и 4850±1000 лет назад, соответственно, и
— семь образцов культуры шнуровой керамики имели мтДНК Т2a1a, T2a1 и U5b1b (4400±400 лет назад), U5b2c (4700±200 лет назад), J1c3 (4500±120 лет назад), H5a и H5a1 (4460±100 лет назад).
К сказанному выше о мтДНК можно добавить, что, действительно, мезолитическая Европа в основном имела гаплогруппу U, что подтверждается добавлением образцов культуры ямочно-гребенчатой керамики, но то, что U – это уже было давно известно из ранних публикаций по ископаемым ДНК. Как известно и то, что с переходом к культуре шнуровой керамики в европейских мтДНК появляется гаплогруппа Н, а также Т и J. В настоящее время около половины всех европейских мтДНК, как западной, так и восточной Европы (включая Финляндию), представляет собой гаплогруппу Н с многочисленными субкладами.
Можно еще добавить, что скелет из культуры шнуровой керамики в Латвии, с мтДНК U5a1 (датировка 4800±200 лет назад), для которого Y-хромосомную гаплогруппу не определяли, находился в скорченном положении, что характерно для гаплогруппы R1a этой культуры. То, что характерно для R1a, в статье, конечно, не указано. Вместо этого авторы сообщили, что ADMIXTURE анализ показал, что этот образец имеет на диаграмме «голубой компонент, который доминирует во всех других образцах из Латвии» (то есть и тех, что с гаплогруппой R1b), «но имеет также и зеленый компонент, который достигает максимума в образцах охотников-собирателей с Кавказа», и далее, что «предковость, относящаяся к охотникам-собирателям с Кавказа, была ранее постулирована (вот так! – ААК), что она прибыла в Европу со скотоводами из Понтийских степей». И дальше авторы продолжают, «на голубом глазу», что в данном образце шнуровой керамики с датировкой 4800±200 лет назад «присутствует степной компонент». Попросту говоря, характерная для таких статей абракадабра. То, что написано, вообще ни к чему не применимо, ни к R1a, ни к R1b, ни для любой другой гаплогруппы. Но понятно, откуда это взялось. Это все эхо статей «широкогеномных попгенетиков» 2015 года (Haak и др., Allentoft и др.), которые направили насельников ямной культуры (R1b) в Европу для создания культуры шнуровой керамики (R1a), что есть абсурд, как уже прокомментировано в предыдущем разделе. И вот теперь авторы очередной работы это всё переписывают.
Как читатель давно сообразил, столь подробные описания абракадабры, порождаемой (или, как написали сами авторы, постулируемой) «широкогеномными» авторами, призваны показать, насколько проще и надежнее выводы ДНК-генеалогии на основе тех же экспериментальных данных. Конечно, «широкогеномные» материалы имеют несравненно более широкий захват, в них анализируются (правда, обычно очень формально) сотни тысяч и миллионы снипов по всем хромосомам, мужчин и женщин, но этот «захват» обычно губит всю работу. Губит потому, что анализ формальный, с массой допущений и приближений, интерпретации совершенно произвольные, альтернативные варианты практически не рассматриваются, и понятно, почему – их в рамках методологии авторов бесконечное множество. А главным образом потому, что не хватает стержня, который позволял бы выбирать из бесконечного множества вариантов интерпретаций только те, которые резко сужают варианты выбора. Это и дают данные и выводы ДНК-генеалогии.
Если бы сочетать геномный анализ с подходами ДНК-генеалогии, выводы статей были бы несравненно более определенными и надежными. Но «широкогеномные» от этого уклоняются, и причин для того немало. Какие основные причины – многократно описывалось на Переформате. Это – и отсутствие школы научного мышления, и беспокойство за свою «делянку», и желание продолжать бесконтрольность, тупо гнать материал, и пугало, которое делают из ДНК-генеалогии, а почему делают – см. предыдущее перечисление причин. Немалая роль в создании этого «пугала» принадлежит Балановским, матери и дитя, которые по сути совершенно далеки от науки. Результат – налицо. Большего ущерба своей области науки, который нанесли Балановские, трудно представить. Надеюсь, наука в этом разберется, и скорее рано, чем поздно.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















