
Гаплогруппы митохондриальной ДНК являются таким же мощным инструментом ДНК-генеалогии, как и Y-гаплогруппы, поскольку содержат информацию о прямых предках по материнской линии. Митохондриальная ДНК передается от матери ко всем ее детям, от дочерей к внукам. Таким образом, мы не можем унаследовать митохондриальную ДНК от отца, только от матери. Митохондриальная Ева – имя, которое дали молекулярные биологи женщине – последнему общему предку по материнской линии всех людей, живущих на планете. Также принято считать, что все современные европейцы являются потомками всего лишь 7 женщин – «дочерей Евы» – прибывших в Европу в разное время и из разных мест. Тест на определение митотипа могут заказать и женщины, и мужчины, желающие узнать свою материнскую ДНК-генеалогическую линию. Интерпретацию результатов базового мтДНК-теста проводят партнёры Академии ДНК-генеалогии – эксперты Deep Dive Group – российской консалтинговой группы в сфере биотехнологий и исторической генетики, при этом историческая часть исследований осуществляется, при необходимости, совместно с международным научным журналом «Исторический формат».

Классическим трудом по исторической генетике женской митохондриальной ДНК выступает «Семь дочерей Евы». Автор этой книги Брайан Сайкс дал условные имена этим семи прародительницам, приняв за первую букву имени обозначение соответствующих вариаций мтДНК, а именно: Хелена (Н), Жасмин (J), Урсула (U), Катрин (K), Тара (Т), Ксения (Х) и Вельда (V). Английскому ученому удалось приблизительно указать эпоху жизни и локализовать родину каждой из семи женщин на карте Европы (которые, впрочем, уточняются) и создать их лирические описания. Они звучат примерно так.
Старшая из всех Урсула, жила около 45 тысяч лет назад в сегодняшней Греции. Она отличалась стройностью и грацией. Условная Ксения разводила огонь и нянчила детей на южных берегах Черного моря, в лесах, изобиловавших волками и медведями. Было это 25 тысяч лет назад. Ксению отличали домовитость и гостеприимство. Дальше следует Тара, жившая 17 тысяч лет назад. Эта женщина, воплощение твердости и непреклонности, родилась в сегодняшней Тоскане, главный город которой – Флоренция. Здесь еще до римлян обитали таинственные этруски, однако, этот исчезнувший народ произошел, скорее всего, не от Тары, ибо он был не индоевропейского корня. Потомки Тары двинулись на север – в будущие германские земли. На север же, в сегодняшнюю Францию, ушли с Пиренейского полуострова многие из тех, кто вел свое происхождение от Вeльды, отличавшейся умением повелевать. Условная Жасмина – родом из современной Сирии, из племени охотников, которых в числе первых осенила гениальная идея – возделывать землю. Жасмина была хрупкой, как цветок, и столь же привлекательной. Екатерина отличалась искренностью, прямотой и открытостью. Она и ее род кормились преимущественно рыболовством у северных берегов Адриатического моря, в районе сегодняшних Венеции и Триеста. Было это около десяти тысяч лет назад. А больше всего среди нас потомков Елены, в характере которой преобладали веселость, ясность и простота. Елена – родом с отрогов Пиренейских гор, из племени охотников, селившихся примерно в тех местах, где теперь живут каталонцы.
Митохондриальная ДНК присутствует во всех клетках организма. Она находится даже в тех клетках, в которых отсутствует ядро (тромбоциты, эритроциты, клетки стержня волос и т.д.). Поэтому для получения мтДНК хорошо подходят любые ткани организма: кости, зубы, кровь, фрагменты скелетированных трупов. Все результаты тестирования мтДНК сравнивают с так называемой «Стандартной кембриджской последовательностью» (CRS) – первой последовательностью мтДНК, которая была расшифрована в 1981 году в Кембридже («последовательность Андерсена»). К настоящему времени все мутации в анализируемой последовательности отсчитывают от CRS, с которой сравнивают нуклеотидную последовательность исследуемой мтДНК и устанавливают ее генетический профиль. Впрочем, несколько лет назад была предложена уточненная референсная последовательность (RSRS).
Брайан Сайкс работал, прежде всего, с европейскими гаплогруппами. На самом деле митохондриальных гаплогрупп выявлено гораздо больше, причем такие гаплогруппы, как A, C, W, I, D не были описаны Сайксом как мало встречающиеся в Европе, поскольку основные центры их распространения – центральная Азия. Мы же, как сообщество, ориентированное на Евразию, будем вводить в рассмотрение и основные гаплогруппы Азии, и говорить не о 7, а о 12 главных дочерях Евы. В наших интерпретациях шести из семи европейским дочерям Евы будут даны славянские имена, за мито гаплогруппой J будет сохранено ближневосточное имя Ясмина, а остальным пяти – имена, принятые в среде тюркоязычных народов
Семь дочерей Евы Б. Сайкса:
H — Helena, Елена
J — Jasmine, Ясмина
U — Ursula, Юлия
K — Katrin, Катерина
T — Tara, Татьяна
X — Xenia, Ксения
V — Velda, Вера
Остальные пять центрально-азиатских дочерей Евы:
A — Айгуль
С — Чулпан
I — Ильмира
D — Диляра
W — Венера
Теоретически, для точного определения мито-гаплогрупп должно изучаться отклонение мутаций генома по всей rCRS или RSRS. Однако критически важная часть информации может быть извлечена из гипервариабельных сегментов HVR1 и HVR2. Гипервариабельный сегмент (HVR) представляет собой область или регион части ядерной ДНК или D-петля в мтДНК, в которой пары оснований из нуклеотидов повторяются (в случае с ядерной ДНК) или заменены (в случае с мтДНК), при этом мутации происходят с наибольшей по сравнению с другими частями генома скоростью. HVR1 считают «низким разрешением» сегмента ДНК или первоначальным и общим, а HVR2 – более «высоким» и уточняющим.
В зарубежной практике существует большое количество различных исследовательских фирм и сервисов анализа мтДНК, которые, однако, являются достаточно дорогостоящими (по всем rCRS и RSRS). В России в настоящее время также запускаются процедуры массового секвенирования, что важно для программы импортозамещения в области биотехнологий. Поскольку эти процедуры находятся на этапе становления и развития, на сегодня собственная база массового тестирования лишь создается и будет создана в ближайшие месяцы. Но уже сейчас доступен «лайт-тест» – определение митотипа митохондриальной ДНК (HVR1 15990-16431, HVR2 00029-00408) по rCRS, который захватывает не полные сегменты HVR1 и HVR2 и далеко не всю кодирующую область, ссылка. Однако это позволяет предсказать мито-гаплогруппу с высокой вероятностью. Более того, выявленные мутации позволяют уточнить предсказанные данные путем анализа накопленных данных в публичных базах, и усилить вывод по гаплогруппе. Для тех же, кто закажет ещё и Y-тест, московская лаборатория ДНК-генеалогии предлагает существенные скидки.
Рассмотрим пример 1. Результаты лайт-теста по rCRS выдали следующие параметры, причем мутации указали на гаплогруппу V (Вера).
HVR I – 16298C; HVR II – 72C 263G 302.1C 310.1C
Данный случай легко поддается интерпретации. По состоянию на середину августа 2016 г. в базе FTDNA есть единственный образец с редкой и нетипичной для V мутацией 310С, который экспертами FTDNA был отнесен к субкладу V2b, ссылка.

Анализ накопленных фактов по истории данной мито-гаплогруппы делает такой вариант правдоподобным. Гаплогруппа «Вера» появилась в палеолите в одном из ледниковых убежищ Западной Европы, предположительно во франко-кантабрийском регионе. При этом ключевым событием истории этой мито-гаплогруппы является ее миграция на Северо-восток вплоть до Скандинавии и до Волго-уральского региона. Данная миграция не была многочисленной, и поэтому потомков Веры в Европе проживает незначительное количество. Заслуживает внимание сравнительно высокая концентрация этой мито-гаплогруппы у басков и саамов. У последних имеется аномально большой процент гаплогруппы V – в среднем 42%, варьируясь от 33-37% популяции финских и норвежских саамов до 68% популяции шведских саамов (у саамов России эта митохондриальная гаплогруппа отсутствует). Соседи саамов – шведы, норвежцы, карелы и финны не обнаруживают большого процента распространения данной гаплогруппы (от 3% у шведов и норвежцев до 6% и более у финнов и карелов). У русских процент носителей женской гаплогруппы V больше, чем у германоязычных скандинавов, но меньше, чем у финноязычных народов – 4% популяции. Это может навести на определенные выводы о появлении и распространении данного палеосубклада, но прежде всего, следует отметить, что из всех европейских народов (помимо саамов) в более чем 10% случаев гаплогруппа V встречается лишь у басков (12,4%) и у марийцев (10,2%).
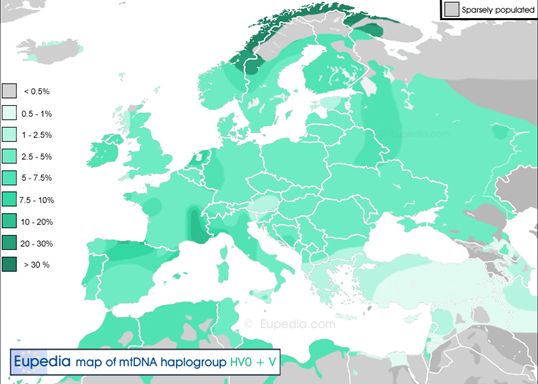
Любопытно также, что у испанцев и португальцев процент гаплогруппы невелик (3-4%), а у мордвы, коми и удмуртов (которые родственны в языковом отношении) еще менее значителен, в последних двух случаях менее 1%. Следовательно, мы имеем дело с определенной миграцией, слоем населения Древней Европы, перекрытым более поздними слоями, который как бы «законсервировался» в ряде обособленных регионов (в т.ч. в Сардинии – 6%).
Также существенным аргументом в пользу появления mtDNA гаплогруппы V в европейских горных и тундровых приледниковых районах является то обстоятельство, что она коррелирует с выявленной предрасположенностью ее носителей к повышенной морозоустойчивости. Это позволяет видеть в гаплогруппе V результат мутации – приспособления к суровым условиям Европы в связи с миграцией на Север. Другие пути расселения мезолитических носителей гаплогруппы V обнаруживаем в неолитические времена на острове Готланд и в линейно-ленточной культуре Германии и в ряде культур неолита и бронзового века Европы. Наиболее хорошо описанный в сообществе палеогенетиков кластер субклада V2 находится на Британских островах, следовательно, возможно, он был среди участников миграции носителей V на Север. Так как носители этой миграции дошли до Волго-Уральского региона, то носители V2 могли оказаться и на Русской равнине. Действительно, они там есть.
Рассмотрим пример 2. Он относится к анализу палеоДНК. Одно из захоронений Южного Оленьего Острова (ссылка), а именно UzOO 77 (5500 лет до н.э.) диагностировано, как имеющее мито-гаплогруппу H («Елена»). При этом отклонения HVR1 от rCRS обозначены как 16235G, 16311C, 16362C.
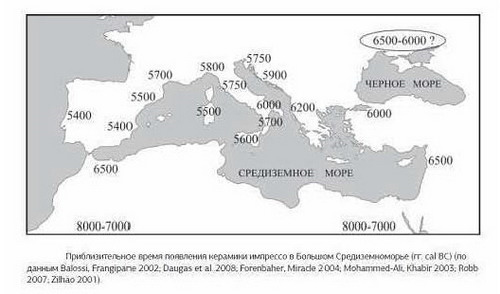
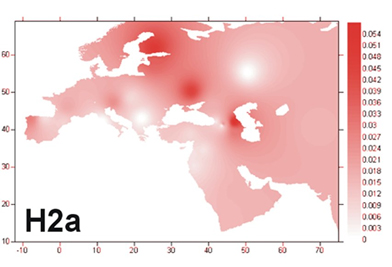
Анализ баз данных может несколько уточнить данный вывод и конкретизировать субклад. В базе данных FTNDA (проект H & HV) был проведен поиск людей, имеющих довольно редкую для H мутацию 16235G. Как показывают результаты анализа базы данных, 73 человека имеют мутацию 16235G. Из них 35 человек (48%) принадлежат к субкладу H2a. Анализ известных на сегодня палеоДНК показывает, что такое положение дел –закономерно. На уровне мезолита-неолита находки H2 не заходят западнее центральной Европы, при этом они обнаруживаются и в других местах находок древней R1a – прото-фатьяновской культуре (жижицкая, 2500 до н.э.), Кум-Тепе (Турция, 4700 до н.э.), Арени (Армения, 4330-3060 до н.э.), Смядово и Виноградное (Причерноморье, 4500-3000 до н.э.), культура воронковидных кубков (Германия, 3400-3000 до н.э.). Известна находка и в культуре Ремеделло (Италия), данная культура связана с более восточной Вучедольской (3500-3000) до н.э.
Женская гаплогруппа H (Елена), предположительно, возникла в Западной Азии около 30 тысяч лет назад, прибыла в Европу около 20-25 тысяч лет назад и распространилась на юго-запад континента вплоть до франко-кантабрийского региона. Но можно говорить о сохранении присутствия H на Ближнем Востоке и Кавказе в мезолите-неолите (Kotias (Грузия) – H13с, Tell Halula (Сирия) – H, Halaf (Турция) – предположительно H3a). То есть первоначальной областью распространения H могла стать широкая полоса от Кавказа и до Франко-Кантабрии.
Выявленные находки H2 укладываются в определенную систему. Учитывая появление этого субклада на севере и востоке Европы уже в мезо-неолите, в широкой полосе от Оленьего Острова, в Причерноморье и до Турции и Армении, и учитывая сохранения южного очага H, позволяет ставить вопрос о миграции с юга. В качестве логичной гипотезы о появлении этого гаплотипа можно считать версию о проникновении в Северную и Восточную Европу морским путем со Средиземноморского бассейна носителей культуры кардиальной керамики. Эта версия позволяет связать появление носителей H2 (и H1) с миграциями (либо иными культурно-археологическими связями) комплекса кардиальной керамики, чьи представители могли передвигаться вдоль побережья на лодках. С причерноморскими носителями кардиальной керамики могли быть связаны носители гребенчатой керамики, след которых просматривается и в процессе сложения фатьяновской культуры, и хвалынской культуры (через самарскую) и, очень возможно, карельской культуры сперрингс (куда можно отнести и Южный Олений Остров, хотя распространена пока и точка зрения о докерамическом характере этого могильника). На Южном Оленьем Острове и в хвалынской культуре выявлены архаические варианты R1a, то есть на этапе неолита носители архаичных субкладов R1a и H2 проживали на одной территории.
Это позволяет считать гаплогруппу H2 исконной для Русской равнины (наряду с U4, U5, H1). Более давние связи уводят корни H2 в Причерноморье и Кавказ. Наличие на Южном Оленьем Острове палеоДНК с Y-гаплогруппой J, а также повышенная концентрация Y-гаплогрупп J2b в Волжском регионе и у саамов, позволяют говорить о возможных миграциях, которые могли связать Турцию и Армению, с одной стороны, и Карелию, с другой, через Причерноморский регион.
Про митохондриальную ДНК, основные определения и исторические отнесения можно еще прочитать в книге А.А. Клёсова «Происхождение славян» (М., 2013, глава 12) и в недавней книге А.А. Клёсова «Ваша ДНК-генеалогия. Узнай свой род» (М., 2016, стр. 26-29). Заказать базовый мтДНК-тест и персональную интерпретацию результатов можно в московской Лаборатории ДНК-генеалогии по этой ссылке.

















