
Перед вылетом в Москву в июне этого года я получил приглашение с механико-математического факультета МГУ прочитать лекцию под названием «Расчетные методы в ДНК-генеалогии». Сначала со мной по этому поводу связался кандидат физ-мат. наук, доцент Г.В. Носовский, известный как автор книг по новохронологии, опубликованных совместно с академиком А.Т. Фоменко. Я поначалу отказался, сказав, что не могу допустить, чтобы мое имя, как и имя Академии ДНК-генеалогии, ассоциировали с новохронологией. Но Глеб Владимирович меня заверил, что «вопросы Новой хронологии не обсуждаются. Впрочем, как и старой (скалигеровской). Однако отсылки к любым консенсусам не приветствуются, поскольку нам, как математикам, они чужды». Последнее положение на простом русском языке означает, что никакое знание, по которому есть согласие между учеными или любыми заинтересованными сторонами, лучше не приводить и на него не опираться. Оно будет оспорено независимо от его очевидности (или кажущейся очевидности), и знание будет вырабатываться в ходе дискуссии по материалам лекции. Или доклада, если его назвать так. Было добавлено, что аудиторию интересуют подробности кинетической модели ДНК-генеалогии. Опять переводя на простой русский язык – как число мутаций в Y-хромосоме переводится в показатели времени, то есть в датировки исторических событий, используя константы скоростей мутаций. По-другому говоря, как число мутаций в Y-хромосоме связано с хронологией древних событий.

Стало понятно, что коллизии с новохронологией не избежать, потому что датировки в ДНК-генеалогии основываются на обычной, общепринятой хронологии. И это, на мой взгляд, совершенно правильно. Например, что Великая Отечественная война началась 75 лет назад; что Октябрьская революция произошла 99 лет назад; что общий предок раввинической семьи Горовицев (группы ашкеназийских евреев гаплогруппы R1a) жил в 1507-1572 гг., то есть 444-509 лет назад; что Джон, Лорд Островов, прямой потомок шотландского короля Сомерледа, умер в 1386 году, то есть 630 лет назад; что арии прибыли в Индию примерно 3500 лет назад; что носители гаплогруппы R1a в синташтинской культуре имеют археологическую датировку примерно 4000 лет назад; что общий предок арабов и евреев в гаплогруппах J1, J2 и R1a, которого в Библии, Торе и Коране называют Авраам, жил примерно 4000 лет назад (это – наши данные); что первые представители Homo sapiens, человека разумного, жили примерно 220 тысяч лет назад (наши данные, подтвержденные расчетами по снип-мутациям), а общий предок шимпанзе и человека жил примерно 4.2 миллионов лет назад (наши расчетные данные по мутациям в Y-хромосомах человека и шимпанзе). Но все эти датировки получены в рамках «консенсуса», то есть общей договоренности о шкале времен, а отсылки к консенсусу, как меня поставили в известность, приветствоваться не будут. Ну что же, это даже интересно. Лучше я буду оппонировать новохронологии, чем выступать в согласии. Поэтому я согласился на лекцию…
Первый же слайд, на котором упоминалось время, тут же вызвал вопрос – откуда появилась скорость снип-мутаций, равная 0.8х10-9 на нуклеотид в год. Вопрос совершенно разумный, особенно если понимать, что его задал новохронолог, у которого концепция времени другая. А именно, что измерения времени другими (не новохронологами) неверные. Поэтому или новохронолог должен убедиться, что они, нормальные измерения, неверные, потому и консенсус в этом отношении не приветствуется, или новохронолог должен признать, что измерения верные, но тогда можно выкинуть все книги по новохронологии, посыпать голову пеплом, и признать, что они, новохронологи, дурачили читателей на протяжении последних тридцати лет или около того. Догадайтесь с одного раза, за что будет биться новохронолог, причем биться до последнего. Да, Г.В. Носовский свое обещание выполнил, слово «новохронология» в ходе дискуссии им произнесено не было, но оно присутствовало в каждом вопросе и комментарии. Другого, собственно, и ожидать не приходилось.

К явному неудовлетворению Г.В. Носовского расчет скорости снип-мутаций на приведенном слайде проводился на основании обычного, «консенсусного» времени, причем по археологическим данным. При словах «археологические данные» Г.В. Носовский иронически-скептически махнул рукой, и было ясно, что эти датировки он не приемлет. Поясним, как эти скорости были получены, причем для сокращения изложения не будем воспроизводить рассуждения авторов цитируемых статей, и используемые ими поправки, допущения и приближения. Покажем суть. Остановимся только на исследованиях последних трех лет. Желающие провести анализ деталей могут обратиться к самим публикациям, на которые мы дадим ссылки.
Итак, в одном исследовании (Poznik и др., Science 341, 562-565, 2013), изучая у двух индейцев племени майя фрагмент Y-хромосомы размером 9.99 миллионов пар оснований, нашли в нем 120 и 126 мутаций, соответственно. Принимая, что заселение Америки произошло 15 тысяч лет назад, они получили, что скорость появления мутаций в Y-хромосоме составляет 120/(9.99×106)/(15×103) = 0.80×10-9 и 126/(9.99×109)/(15×103) = 0.84×10-9, или в среднем 0.82х10-9 мутаций на пару нуклеотидных оснований в год. В другом исследовании (Fu и др., Nature 514, 445-450, 2014) использованы данные по полному геному древнего Усть-Ишимского человека, жившего примерно 45 тыс. лет назад в Западной Сибири. Проанализировав область Y-хромосомы протяженностью 1.86 милионов нуклеотидных пар оснований, авторы обнаружили там 64 мутации, и получили 64/(1.86×106)/(45×103) = 0.76×10-9 мутаций в год на пару нуклеотидных оснований. В третьем исследовании (Адамов и др., Russ. J. Genet. Geneal. 7, 46-67, 2015) авторы использовали опубликованные геномные данные для костных останков человека, жившего в Америке более 12 тысяч лет назад, для которого ветви определенных ДНК-линий разошлись 15621 год назад, и нашли 392 мутации от точки расхождения во фрагментах Y-хромосомы общим размером 30.65 миллионов нуклеотидных пар. Это дало 392/(30.65×106)/(15.621×103) = 0.82×10-9 мутаций в год на пару нуклеотидных оснований.
Принимая скорость снип-мутаций за 0.82×10-9, получаем, что одна мутация во фрагменте Y-хромосомы размером 8.47 миллионов нуклеотидных пар (этот размер фрагмента принят за референсный, то есть стандартный) происходит раз в 8.47х106 х 0.8х10-9 = 144 года. Иначе говоря, если в Y-хромосоме группы людей (или отдельного мужчины) между снипом, маркирующим определенную гаплогруппу, и настоящим временем насчитывается 100 снипов, то эта гаплогруппа образовалась 14400 лет назад. Естественно, с определенной погрешностью, которые вычисляются в каждом конкретном случае.
Действительно, мы видим причину неудовлетворенности новохронолога, потому что здесь использованы археологические датировки 15 тысяч лет, 45 тысяч лет, и 15 с лишним тысяч лет назад. Новохронолог в них не верит, но это, конечно, его личное дело. Мы же видим согласование всех трех скоростей мутаций, или, правильнее, констант скоростей мутаций, потому что они не зависят от абсолютных датировок. Справедливости ради надо сказать, что есть и другие оценки констант скоростей снип-мутаций, которые несколько отличаются от приведенных выше, но в каждом случае ясно, в чем причина различий. Это или неудовлетворительное качество секвенирования Y-хромосомы (то есть определения протяженности нуклеотидных фрагментов и/или числа мутаций в них), или работа не с Y-хромосомами, а с аутосомами, то есть с другими хромосомами ДНК, и так далее. Приведенные выше константы скоростей мутаций дают наиболее согласованные и воспроизводимые датировки по снип-мутациям. Более того, как будет показано, получаемые датировки обычно согласуются и с датировками, получаемыми и по мутациям в гаплотипах, то есть независимыми методами и с другой методологией.
Что касается стир-мутаций, упомянутых на слайде выше, то поскольку в 111-маркерном гаплотипе константа скорости стир-мутации равна 0.198 мутаций на гаплотип за 25 лет (условное поколение), то мутации происходят раз в 1/0.198 = 5 условных поколений, или 125 лет. Это, конечно, чистая случайность, что скорости (или частоты, что по сути синонимы в данном случае) снип-мутаций (раз в 144 года) и стир-мутаций (раз в 125 лет) близки друг другу, но это представляет определенные удобства в расчетах. На самом деле первая величина относится к определенному фрагменту Y-хромосомы (8.47 миллионов нуклеотидных пар), а вторая – к гаплотипу определенной протяженности (111-маркерный), и обе величины можно менять как угодно в любом эксперименте. Взяли бы фрагмент Y-хромосомы размером в 9.76 милионов нуклеотидов, получили бы, что снип-мутации происходят раз в 125 лет, то есть совпали бы со средней скоростью стир-мутаций в 111-маркерном гаплотипе.
Вернемся к лекции. Итак, первая коллизия с новохронологом уже произошла, обычные датировки он не принимает, считает, что обычная (или «консенсусная») шкала времени неправильная. Правильная это та, при который Иисус Христос родился в 1090 году, и он же есть Юрий Долгорукий, что Чингиз-хан вторгся в Среднюю Азию в 1350 году, и был он русским, Георгием Победоносцем, а арии перешли в Индию тогда же, в средние века. Взяли бы мы те, датировки «новохронологии», тогда всё было бы правильно, комар носа не подточит. Была бы настоящая наука. Правда, Г.В. Носовский именно этого в дискуссии на моей лекции не говорил, он же обещал. Датировки я взял из его книг и обсуждений. А что он говорил – это, повторяю, что нельзя брать обычные, общепринятые датировки, которыми оперируют в книгах, журналах, в школах и институтах-университетах, датировки, которые используют археологи, физики, химики, инженеры и все прочие. Поэтому через каждые полминуты он мою лекцию прерывал, и настаивал на том, что мы должны измерять время независимо, «со стороны», «извне», чтобы не поддаваться «консенсусным» представлениям о шкале времени. Или, еще лучше, чтобы измерять время в ДНК-генеалогии в неких условных единицах, только не в годах. Есть ли такие методы? – спрашивал он.
Из аудитории подсказали, что есть независимые методы, которые основываются не на годах, а на одном поколении, измеряя число мутаций, которые происходят в Y-хромосоме между парой отец-сын. Г.В. Носовский оживился, воскликнув, что, мол, вот оно, вот то, что надо! Я вклинился, и пояснил, что, увы, это не то, что надо, поскольку этот метод в его опубликованном исполнении настолько неточный, что толку от него почти никакого. При этом я поначалу допустил разговорную неточность, сказав, что этот метод «пар отец-сын» показывает другие данные, и это вызвало ликование новохронолога – ага! Значит, дает другие данные – ясно, тогда это показывает, что ваши расчеты основаны на неверных посылках! Значит, ваши датировки неверные! Было занятно наблюдать, как новохронолог ищет любую зацепку, чтобы спасти свою новохронологию, хотя для стороннего наблюдателя это могло выглядеть совершенно кошерно – всё разумно, математик ищет истину, которую ему не дают. Мешают, вводят его в заблуждение.

Пришлось объяснить, что там не данные другие, а настолько неточные, что могут иметь только сугубо концептуальное значение, а именно, показывают, что мутации в ходе одного поколения, между отцом и сыном, происходят, и происходят настолько редко, что в большинстве случаев просто не определяются, а когда определяются, то статистики там нет никакой. Пример – в одном случае в некоем эксперименте, повторив измерения сто раз, получили результат 5.0±0.2, а в другом, повторив три раза, получили 4±3. Это как, другие данные? Концептуально, так сказать, то же самое, но второй вариант по сравнению с первым практически бесполезен. Вот так, вторым вариантом, и получают «скорости мутации» по парам отец-сын. Они не противоречат тем, что получены первым вариантом, и это все полезное, что от них можно получить.
Приведем конкретные данные. В статье Ballantyne и др. (Amer. J. Hum. Genet. 87, 341-353, 2010) изучали переходы мутаций от отца к сыну в 187 маркерах Y-хромосомы, при этом измерения проводили на 1400-1800 пар отец-сын (это число было разным для разных маркеров). В эти 187 маркеров входили большинство (но не все) из 111 «штатных» маркеров протяженных гаплотипов. То есть сама постановка эксперимента уже делала невозможной применение всех 111 маркеров для расчетов времен жизни общих предков по гаплотипам, поскольку маркеры были с многочисленными пропусками. На самом деле там было еще хуже: любая панель маркеров (12-, 25-, 37-, 67-, 111-маркерная) имела пробелы по изучавшимся маркерам, ни одна не была полной, ни одна не могла быть применена для расчетов на основе мутаций в гаплотипах по этим панелям. Ну да ладно, хотя и неудобно, но можно было бы сконструировать какую-нибудь гибридную, нестандартную панель, и с ней работать, правда, коллеги бы не поняли, зачем это нужно.
Но настоящая беда была в том, что из 187 изучавшихся маркеров в 67 из них мутаций вообще не было, то есть никаких скоростей мутаций из них было не получить. Из 111 «штатных» маркеров мутаций не было в 24 маркерах, а еще для 56 маркеров данные были совершенно ненадежны. Какая константа скорости для маркера, когда за поколение в маркере прошла всего одна мутация? Как константу будем рассчитывать? А если прошло всего 2-3 мутации, как константу рассчитывать будем? При погрешности в 70-100%? Пробовали бросить монету три раза, и по результатам рассчитать вероятность выпадения орла или решки?
Коротко говоря, эти данные по парам отец-сын для расчетов вообще непригодны. Возьмем, например, опять те же цитируемые результаты. В маркере DYS393, весьма «медленном», у 1750 пар отец-сын прошли три мутации, а в маркере DYS390, намного более «быстром», у 1758 пар прошли две (!) мутации, то есть маркер оказался как бы более медленный, что есть нонсенс. Любой, кто работает с гаплотипами, знает, что в DYS390 проходит больше мутаций, чем в DYS393. Вот пример – среди 3466 гаплотипов субклада R1b-L21 в маркере DYS393 от общего предка прошло 232 мутации, а в маркере DYS390 – 1164 мутации, в пять раз больше. А по парам отец-сын у Ballantyne – наоборот, 3 и 2 мутации, соответственно. Можно на основании расчетов по таким данным на что-то надеяться? Нет, конечно, и понятно, почему – статистика не та. Мало для расчетов 1750 пар отец-сын, очень мало.
Далее, если даже взять 100 тысяч пар отец-сын, то те маркеры, у которых константа скорости мутации меньше чем 0.0001 мутаций на маркер на поколение, все равно покажут плохую статистику, у них в среднем будет наблюдаться меньше, чем 10 мутаций на поколение для всех этих 100 тысяч пар. То есть погрешность определения константы скорости мутации для них в лучшем случае в среднем будет ±32%. Но это только для тех, у которых константа скорости 0.0001. А таких, с константами 0.0001 и меньше, в 111-маркерной панели девять маркеров, из них пять – в 67-маркерной панели (показана константа скорости на маркер за условное поколение в 25 лет):
DYS472 0.000008
DYS436 0.000040
DYS425 0.000042
DYS568 0.000050
DYS490 0.000070
DYS426 0.000090
DYS455 0.000100
DYS632 0.000100
DYS494 0.000100
Кстати, для всех этих девяти маркеров в серии из 1500-1800 пар отец-сын (Ballantyne, 2010) не наблюдалось ни одной мутации. То есть их данные опять неприменимы для расчетов. А для DYS472 и 100 тысяч пар-отец-сын недостаточно и для наблюдения хотя бы одной мутации. Там маркер мутирует в среднем раз в 125 тысяч поколений, то есть раз на 125 тысяч пар отец-сын. В общем, меньше чем полумиллионом пар не обойтись, лучше – миллион пар отец-сын. Это, конечно, практически нереально.
Есть еще проблема с парами отец-сын. В них константы получаются в расчете на одно поколение. А в исторических науках на поколение не считают, нужно пересчитывать в года. А как? Популяционные генетики эту проблему так и не решили. Так что и в этом отношении пары отец-сын практически бесполезны. Еще проблема – таких опытов в парах отец-сын проведено несколько, и результаты в них, как правило, не совпадают друг с другом. Причина все та же – статистика недостаточна. Приведем несколько примеров, сопоставив цитируемую выше работу со сводной таблицей (Burgarella и др., Eur. J. Hum. Genet. 19, 70-75, 2011), в которую сведены данные, полученные суммарно более чем с 12 тысячами пар отец-сын в разных экспериментах. В первой цитируемой работе в маркере DYS393 три мутации на 1750 пар (что в дальнейшем будем обозначать как 3/1750), во второй – 13/12576 (при пропорциональном пересчете 1.8/1750), отношение в скоростях составляет (3.0/1.8) 167%.
В маркере DYS390, 2/1758 и 30/14131, соответственно, разночтение в скоростях 182%.
В маркере DYS391, константы скоростей мутаций оценены в диапазоне 0.00126-0.00665 (Ballantyne) и 0.00198-0.00372 (Burgarella). Формально диапазоны перекрываются, но вести расчеты с таким широким диапазоном неопределенности смысла не имеет. По нашим независимым данным (таблица была показана на лекции, и приведена ниже) величина константы скорости мутации для DYS391 равна 0.00220 мутаций на маркер на 25 лет, то есть в том же диапазоне.
DYS385a,b – данных у Burgarella нет, у Ballantyne дается диапазон 0.000624-0.00506 для DYS385a, то есть 8-кратный диапазон неопределенности, и 0.00175-0.00809 для DYS385b, то есть диапазон неопределенности в 4.6 раз. По нашим независимым данным величина константы скорости мутации для DYS385a и DYS385b равна 0.00280 и 0.00360 мутаций на маркер на 25 лет, соответственно, то есть опять попадает в широкий диапазон результатов прямых экспериментов. То есть не противоречит, но толку от того мало.
Для DYS398-2 у Ballantyne (прошло 6 мутаций за поколение) получилась скорость мутаций в диапазоне между 1.6 и 7.5 на 10-3, у нас 2.31х10-3.
Для DYS391 (5 мутаций) у Ballantyne между 1.26 и 6.65 на 10-3, у нас 2.2х10-3.
Для DYS439 (6 мутаций) у Ballantyne между 1.63 и 7.54 на 10-3, у нас 3х10-3.
Для DYS456 (8 мутаций) у Ballantyne между 2.35 и 8.97 на 10-3, у нас 4.32х10-3.
Для DYS458 (14 мутаций) у Ballantyne между 4.80 и 13.4 на 10-3, у нас 6.2х10-3.
Для DYS464 (12 мутаций, суммарная величина по четырем мультимаркерным локусам) у Ballantyne между 3.96 и 12 на 10-3, у нас 6.0х10-3.
Перечень можно продолжить, это продолжение с детальным обсуждением дано в недавней книге «Кому мешает ДНК-генеалогия» (Книжный мир, 2016, 848 стр.), но для данного рассмотрения достаточно. Вывод очевидный – результаты независимых прямых экспериментов по определению мутаций в парах отец-сын не противоречат нашим данным, но они, эти результаты, имеют настолько большую погрешность, и во многих случаях искажены вследствие недостаточной статистики, что их даже рассматривать большого смысла не имеет.
Много вопросов со стороны новохронолога было о том, совпадают ли результаты расчетов по разным маркерам, ведь все они должны быть связаны с накоплением мутаций от базового гаплотипа, от общего предка. Иначе говоря, считать по 12-маркерным гаплотипам, 17-маркерным, 25-маркерным, 37-маркерным, 67-маркерным или 111-маркерным, не должно иметь большого значения, они должны согласовываться. Понятно, что чем больше маркеров, тем точнее расчеты, тем меньше погрешность расчетов, но это тогда, если они согласуются между собой. Вопрос правильный, ответы были даны на разных примерах.
Пример один – длинная серия из 3466 гаплотипов в 111-маркерном формате, всего 384726 аллелей. Из них можно «выхватывать» гаплотипы более короткие, и повторять расчеты времени до общего предка всей серии гаплотипов. Например, как показано на следующем слайде.

Аналогично, при проведении расчетов, когда жил общий предок арабов и евреев гаплогруппы J1, опять получились взаимно согласующиеся данные.

Для слушателей такие согласования разных панелей маркеров были, видимо, удивительны, ведь в каждом случае использовалась другая константа скоростей мутаций, «калиброванная» под каждую панель. Например, для 6-маркерных гаплотипов она равна 0.0074 мутаций на гаплотип на 25 лет, для 12-маркерных 0.02, для 25-маркерных 0.046, для 37-маркерных 0.09, для 67-маркерных 0.12, для 111-маркерных 0.198 мутаций на гаплотип на 25 лет. Но для нас это было совершенно закономерно, это же ведь отрабатывалось на сотнях и сотнях примеров, в том числе и документальной генеалогии.
Был показан и пример с документальной генеалогией. Приведенные пять ашкеназийских евреев – это знаменитая в еврейской среде линия Горовицев, пражских раввинов.

Здесь «расчеты на ККК» означает расчеты с помощью калькулятора Килина-Клёсова, отсюда и ККК. Калькулятор считает более тщательно, чем ведутся расчеты вручную – он не округляет, вводит более точные поправки на возвратные мутации (при которых мутация возвращается в исходное, немутированное состояние, как будто бы мутации не было). Как видно, и здесь расчеты по гаплотипам разной протяженности дали практически одинаковые результаты, причем в разумном согласии с документальной генеалогией.
Но вопросы от Г.В. Носовского не прекращались. Видимо, он никак не мог примириться с тем, что «нормальная» хронология работает, и работает успешно. Причем работает «вдоль и поперек» и временной шкалы, и разных панелей гаплотипов, и разных серий гаплотипов, от пяти (см. выше) до тысяч гаплотипов, и при расчетах разными способами – линейным (в которых считается число мутаций от базового гаплотипа, который обычно равен предковому), квадратичным (где предковый гаплотип не нужен, мутации считаются «всех от всех», попарно), и логарифмическим (в которых мутации вообще не считаются, считается число сохранившихся, не мутированных гаплотипов). Но Г.В. Носовский неустанно искал, в чем все-таки можно усомниться. А кто ищет, тот всегда найдет. Например, астрономы смотрят на Луну и звезды в телескоп, а в микроскоп почему-то не смотрят. Это непорядок, так наука не делается. Надо смотреть и сверять.
Так, в общем, получилось и у нас, в ходе дискуссии. В ходе изложения я рассказывал, что маркеры – это в нашем случае участки в Y-хромосоме, каждый состоит из десятков и сотен нуклеотидов, и в каждом маркере определенный нуклеотидный фрагмент (часто тринуклеотид или тетрануклеотид, бывает и больше) повторяется подряд несколько раз, обычно от пяти раз до 45. Потому их еще называют «тандемными». Мутация в данном случае – это изменение числа повторов «тандема». Было, например, повторение тринуклеотида 16 раз, и так, прогонами по 16, биологическая машина организма копировала этот маркер тысячелетиями. И вдруг – ошибка, сбой копирующей машины, скопировалось от отца к сыну не 16 раз, как завещали предки, а 15 или 17 раз. Это и есть мутация. К действию радиации или других внешних факторов эти мутации не имеют никакого отношения, потому скорости мутаций (или, более правильно, константы скорости мутации), одинаковы у людей по всей планете, независимо от их родовой принадлежности, антропологических особенностей, расы и прочего. Нет никаких данных, что скорости мутации изменяются ни по регионам, ни во времени. Появятся такие данные – рассмотрим, но пока не было, хотя мутации считали по тысячам и тысячам гаплотипов по всему миру, и если считали, используя выверенные методы, то всегда получали данные в согласии друг с другом.
А поскольку у каждого маркера своя нуклеотидная структура, то их перестройка, это и есть мутация, происходит у одних маркеров относительно быстро, у других медленно. Скорости мутации при этом различаются у разных маркеров на несколько порядков величин. Самый «быстрый» маркер (DYS710) мутирует с константой скорости 0.0073 мутаций за 25 лет (условное поколение), то есть мутирует раз в 137 условных поколений, или раз в 3425 лет. Можно сказать и по-другому – мутация этого маркера происходит раз на 137 рождений мальчиков. А самый медленный (DYS472) мутирует с константой скорости 0.000008 мутаций за 25 лет, то есть раз за 125 тысяч условных поколений, или раз в 3 миллиона 125 тысяч лет. Или же раз на 125 тысяч рождений мальчиков, или раз на 125 тысяч пар отец-сын. Понятно, что при числе в 1700 пар отец-сын вероятность прохождения мутаций всего лишь около 1 процента. Легко посчитать, что самый «быстрый» маркер мутирует со скоростью почти в тысячу раз быстрее, чем самый «медленный» маркер.
Но это не понравилось нашему новохронологу, он это назвал «разбросом» в скоростях, и к этому не раз возвращался, выражая сомнение, что такой «разброс» при усреднении приведет к правильным расчетам. У меня профессия другая, и я с такими «разбросами» постоянно имею дело, как и любой из читателей. И «разбросом» мы это не называем. Например, любая химическая реакция отражает взаимодействия молекул, которые соударяются друг с другом с разной интенсивностью, одни вообще разлетаются при ударе, не сопровождаясь никакой химической реакцией, соударения других приводит к реакции, и все это усредняется по миллионам соударений, и в итоге мы измеряем такие «усредненные» скорости реакций, которые замечательно воспроизводятся от опыта к опыту. Никому эти усреднения не мешают. Аналогично, давление воздуха в автомобильной шине складывается и усредняются по миллионам соударений молекул газов, составляющих воздух, с внутренней поверхностью шины, и ничего, давление постоянно и держится долго, если нет утечки воздуха.
Более близкая аналогия с обсуждаемой ситуацией – это бассейн, из которого ведет много трубочек и труб для слива воды, одни трубы широкие, по ним вода вытекает быстро (аналог высокой скорости мутации), по тонким трубочкам вода вытекает медленно. А в сумме – столько-то кубических литров или кубометров воды в единицу времени. И никому не мешает этот расход воды просуммировать, и то и усреднить. Получим, например, что вытекает на круг (то есть по аналогии на все 111 маркеров гаплотипа) 1200 литров воды за час, вот и хорошо, аналог скорости убывания предковых гаплотипов по всем 111 маркеров, кумулятивно. А можно работать и по всем 111 маркерам, то есть трубочкам и трубам, индивидуально, в сумме должно получиться то же самое, в чем проблема-то? Так что и здесь сомнения новохронолога ни к чему не приводят. А поскольку я работаю в естественных науках, к которым математика не относится, как известно, то у меня давно выработался принцип восприятия сомнений. Сомневаешься – рассчитай сам, и покажи, в чем ошибка. А сидеть на заборе и выражать общие сомнения в том или в этом – это не дело. Для меня – это проблема у «сомневающегося» с этикой в науке.
В ходе доклада затронул я и важную тему расчетов особенно древних событий, приведя ряд примеров по вычислению датировок образования древнейшей гаплогруппы A00 (220 тысяч лет назад), и даже времени расхождения ДНК-линии общего предка современных шимпанзе и человека – 4.2 миллиона лет назад. Вообще при постепенном переходе в расчетах к временам более 10 тысяч лет назад мутации в ряде маркеров, а именно маркерах «быстрых» становятся нестабильными. Доля возвратных мутаций прогрессивно нарастает, аллели начинают бегать взад-вперед, и нарастает «насыщение» системы мутациями. Поэтому при временах событий более 10-20 тысяч лет назад расчеты в ДНК-генеалогии переходят на «медленные» маркеры, и для этого специально разработана панель из 22 самых «медленных» маркеров. Полученные при этом датировки хорошо согласуются с теми, что расчитаны по снип-мутациям (там расчет идет по другому принципу) и с данными антропологов. Никаких вопросов быть не должно, практика – критерий истины.
Однако опять Г.В. Носовский – «а вы по одним быстрым маркерам расчеты вели?». Я отвечаю, что специально нет, потому что это неинтересно. А неинтересно потому, что когда мы ведем расчеты времен жизни недавних общих предков групп гаплотипов, там не нужно убирать медленные маркеры, они и так участия в расчетах не принимают. Смотрите – выше был пример расчетов времени жизни общего предка для пяти ашкеназийских евреев, он жил по данным документальной генеалогии 444-509 лет назад, то же получилось и по расчетам ДНК-генеалогии. А медленные маркеры мутируют раз во много тысячелетий, ясно, что в этом случае в них мутаций не будет, и в игре останутся только быстрые маркеры. Зачем нам их специально выделять из панелей? Медленные маркеры сидят себе тихо, и никому не мешают.
Чтобы быть предметным, можно и конкретные данные привести. На слайде выше (про ашкеназийских раввинов) было указано, что из 555 аллелей (5 гаплотипов по 111 маркеров) только в 17-ти прошли мутации. Посмотрим, что это за маркеры, но уже и так ясно, что это будут самые «быстрые». Так и оказалось. Чтобы не приводить эти 17 маркеров, заметим, что константы скорости мутации в 111-маркерных гаплотипах можно подразделить на три категории – «быстрые», выше 0.001 мутаций на условное поколение на маркер; «средние», от 0.001 до 0.0001; и «медленные», ниже 0.0001 мутаций на условное поколение на маркер. Из тех 17 мутаций 14 оказались в самом «быстром» диапазоне, одна на границе между «быстрым» и «средним», и две – в «среднем» диапазоне. Так что все равно главную роль в недавних ДНК-генеалогиях играют «быстрые» скорости мутаций.
По ходу лекции были кратко рассмотрены константы скоростей мутации для всех 111 маркеров
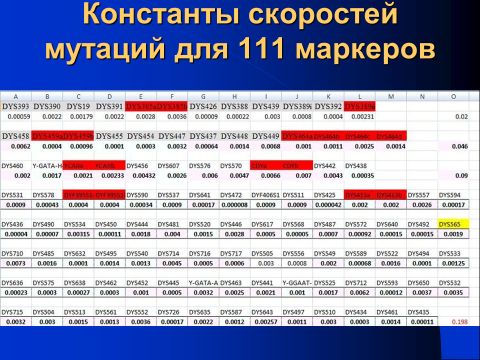
и то, как эти величины были получены, на конкретных примерах «быстрых», «средних» и «медленных» маркеров, как показано на следующих трех слайдах



и много других практических примеров. Коллизии в итоге заглохли, поскольку стороны свои задачи выполнили. Г.В. Носовский показал, что он продолжает скептически относиться к показателям времени, принятым в научном мире и в быту, а я показал, что ДНК-генеалогия не имеет ни малейшего отношения к «новохронологии». Что и требовалось доказать.
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















