
Продолжаем публиковать небольшие фрагменты из книги А.А. Клёсова «Кому мешает ДНК-генеалогия», которая вскоре увидит свет. Книга будет довольно солидной по объёму, около 850 страниц. У читателей сайта и членов клуба Переформат будет эксклюзивная возможность получить эту книгу первыми, следите за обновлениями на Переформате. А пока следующие вопросы и ответы, которые помогают лучше понять ДНК-генеалогию. По этой ссылке первая часть…

Как в Y-хромосоме образуются мутации?
Время от времени при копировании Y-хромосомы в копии проскакивают ошибки. Фермент под названием ДНК-полимераза (на самом деле вместе с целым набором молекулярных инструментов копировальной машины) или просто ошибается и делает некоторые участки ДНК короче или длиннее, удлиняя или сокращая «тандемные повторы» нуклеотидов, или «ремонтирует» повреждённые участки и в ходе «ремонта» удаляет повреждение (эта операция называется «делеция»), или «вшивает» новый нуклеотид (называется «вставка», или «инсерт»). То, что удалось отремонтировать, в мутации не попадает, как будто мутации и не было. Что не удалось — передается сыну при рождении. Если родилась дочь, то Y-хромосома не передается. Если есть только дочери, или детей нет — Y-хромосома терминируется, вместе с мужской наследственной линией.
Мутации, которые интересуют ДНК-генеалогию, бывают двух типов — или изменение числа повторов, тандемов (STR), что описано выше, или «точечные», одно- или несколько-нуклеотидные (SNP), снипы. Оказалось, что многие повторы нуклеотидов находятся у разных людей в одних и тех же участках Y-хромосомы. Эти участки уже специалистами пронумерованы, классифицированы, сведены в списки. Они получили название «маркеры». Набор маркеров, точнее, повторов в них, называется «гаплотип». Уже известных маркеров — больше тысячи. Они, как правило, одни и те же у всех людей на Земле и отличаются друг от друга только числом повторов, что вызвано теми самыми ошибками ферментов (и других элементов биологического копирования) при копировании из поколения в поколение.
Ясно, что «мутации», которые приводят к изменению числа тандемных повторов в Y-хромосоме, не есть те «мутации», о которых все имеют представление как о «поломках в генах», например, под воздействием радиации. Мутации, которые рассматривают в гаплотипах, происходят не в генах и не являются «поломками». Это – ошибки при копировании ДНК. Они имеют весьма сложную архитектуру, и радиацией не вызываются. Это – не поломка, а координированная перестройка.
Что такое «общий предок» выборки носителей гаплотипов, и как мутации могут помочь определить, когда он жил?
Любая группа мужчин, относящихся к одной и той же гаплогруппе и к одному и тому же субкладу, а также к любой выборке гаплотипов, образующей ветвь в пределах одного субклада, являются в каждом случае родственниками, и происходят от одного общего предка. Можно привести и обратное определение – если мутация, которая осуществилась в Y-хромосоме, унаследовалась в ДНК потомков, то все эти потомки происходят от одного общего предка, у которого и произошла данная мутация.
В принципе, в любой гаплогруппе и любом субкладе имеется множество групп людей, которые происходят от их общего предка. Например, у двух родных братьев есть общий предок – их отец. Если к ним добавить дядю, то общий предок всех троих – отец дяди и дедушка братьев. Чем глубже во времени общий предок группы людей, или популяции, в терминах ДНК-генеалогии, тем больше у него потомков. Все потомки наследуют гаплотип их общего предка, но чем дальше во времени отстоит общий предок, тем больше мутаций накапливается в гаплотипах потомков.
Приведем простой пример. Допустим, общий предок жил 2500 лет назад, то есть 100 условных поколений назад (в ДНК-генеалогии за условное поколение принимается 25 лет, и константы скоростей мутации вычисляются в расчете на условное поколение протяженностью 25 лет). Если выявить сто потомков этого общего предка, в их 12-маркерных гаплотипах обнаружится примерно 200 мутаций, причем у 14 человек гаплотипы будут одинаковыми, и все 200 мутаций будут неупорядоченно распределены, в разной степени, между оставшимися 86 потомками. Вот эти одинаковые 14 гаплотипов и представляют собой предковый гаплотип. За прошедшие 100 поколений – чисто статистически – в гаплотипах 86 потомков пройдут мутации, а в гаплотипах 14 потомков мутаций еще не будет. Вот таким образом определяют предковый, или базовый гаплотип.
Проверим представленные выше числа. 200 мутаций в 100 гаплотипах в 12-маркерном формате (константа скорости мутаций в 12-маркерных гаплотипах равна 0.02 мутаций на гаплотип за условное поколение) соответствуют величине 200/100/0.02 = 100 условных поколений, то есть примерно 2500 лет (здесь для простоты не вводится поправка на возвратные мутации, смысл которой будет пояснен в следующих частях). Это – линейный метод расчета времени до общего предка популяции. В то же время согласно логарифмическому методу если из 100 гаплотипов в 12-маркерном формате остались неизменными 14, то это соответствует времени, прошедшему от общего предка популяции, равному [ln(100/14)]/0.02 = 98 условных поколений, то есть примерно тому же времени, рассчитанному с помощью линейного метода.
Из этого примера должно быть понятно, что чем более недавний общий предок, тем большее число идентичных друг другу гаплотипов (или их фрагментов) наблюдается в выборке, и тем надежнее выявляется предковый гаплотип. Если общий предок жил очень давно, то одинаковых гаплотипов в выборке может остаться всего два-три – например, три гаплотипа из сотни 12-маркерных гаплотипов останутся при [ln(100/3)]/0.02 = 175 условных поколений до общего предка, то есть общий предок жил 4375 лет назад (на самом деле больше при учете возвратных мутаций). Когда в серии гаплотипов наблюдаются всего два-три одинаковых гаплотипа, они могут не обязательно быть предковыми, а оказаться одинаковыми случайно, в результате игры неупорядоченных мутаций. Поэтому понятие «предковый» гаплотип лучше не употреблять, за исключением совсем уже явных случаев, и в ДНК-генеалогии вместо «предковый» гаплотип обычно употребляют «базовый».
Итак, общий предок популяции – это предполагаемый носитель «базового», или «предкового» гаплотипа. Время жизни общего предка популяции вычисляется по совокупности гаплотипов его потомков, принимая во внимание число мутаций, накопившееся в популяции, нормированное на гаплотип или на маркер, и константу скорости мутации, или частоту мутации в расчёте на поколение. Расчёты времён жизни общих предков популяций позволяют делать предположения о времени и направлениях миграций популяций в древности, о передвижениях популяций. Поскольку, передвигаясь, популяции оставляли следы в виде материальных признаков, это позволяет проводить интерпретацию археологических данных в отношении носителей этих признаков, их принадлежности к определённым родам и племенам, связанных родственными взаимоотношениями. Аналогично, популяции, передвигаясь, приводили к перемещению языков, диалектов, и определение общих предков и времён их жизни даёт возможность прослеживать динамику языков в отношении соответствующих родов и племён, связанных родственными взаимоотношениями.
Можно ли привести примеры предковых гаплотипов, датируемых десятками и сотнями тысяч лет назад? Можно ли выявить гаплотип общего предка всех мужчин на Земле?
«Общий предок всех людей» – понятие скорее концептуальное, чем буквальное, абсолютное. Чем больше накапливается данных, тем понятнее становится, что «общий предок всех людей» теряется среди переплетений ДНК-генеалогических линий, уходящих на сотни тысяч лет вглубь. Поэтому введенное некоторое время назад понятие «хромосомный Адам» не является научным и никогда не было доказано. Интуитивно, такой общий предок возможен, но он, скорее всего, будет «плавающей мишенью».
Понятие «одного предка» можно рассматривать на разных уровнях сложности. Например, древнейшая африканская линия (гаплогруппа А00) имеет общего предка с гаплогруппой R1a примерно 210 тысяч лет назад, то есть оттуда, из такой глубины тысячелетий расходятся гаплотипы современных носителей гаплогруппы A00 и этнических русских гаплогруппы R1a:
13 11 12 10 11 16 10 8 14 14 8 8 8 9 12 11 12 8 12 12 11 11 (А00)
12 12 11 11 11 11 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a)
Здесь приведены 22 очень «медленных» маркера Y-хромосомы, каждый из которых мутирует раз во многие тысячелетия. Можно подсчитать, что эти гаплотипы разошлись на 26 мутаций, и расчеты, основанные на скорости мутации каждого маркера, показывают, что эти 26 мутаций соответствуют времени жизни их общего предка примерно 210 тысяч лет назад. Но даже беглый взгляд на эти гаплотипы показывает, что ДНК древнейших африканцев и этнических русских, носителей гаплогруппы R1a, не так уж намного различаются, или, скорее, являются принципиально сходными.
Хорошо, а если вместо типичного гаплотипа R1a подставить гаплотип R1b, преобладающей гаплогруппы Западной Европы? Мутации здесь настолько медленные, что в таком формате представления гаплотипа он будет практически одинаков по всей Западной Европе:
11 12 13 11 11 12 11 9 15 16 8 10 8 12 10 12 12 8 12 11 11 12 (R1b)
Мы видим, что при таком разрешении большая часть аллелей (это — числа в гаплотипах) у R1a и R1b одинаковы (совпадают 16 аллелей из 22). Между гаплотипами А00 и R1b — 24 мутации, что снижает возраст общего предка на семь тысяч лет, то есть на 3%. Это — в пределах погрешности расчетов. Только не стоит строить пропорции, там счет нелинейный.
Можно ли считать, что общий предок человечества жил 200-210 тысяч лет назад? Вряд ли, поскольку гаплогруппа А00 была обнаружена всего два года назад, и до этого считали, что общий предок человечества жил 140 тыс. лет назад. Если завтра исследователи найдут новую гаплогруппу, то срок начала человечества опять отодвинется. И он несомненно будет отодвигаться дальше, по мере углубления наших знаний. Уже появилась еще одна реперная точка — это общий предок шимпанзе и человека. У современного шимпанзе из полной хромосомной последовательности удалось определить 16 маркеров Y-хромосомы. Они показаны рядом со значениями аллелей в тех же маркерах Y-хромосомы «усредненного общего предка» человека:
8 15 10 4 5 9 10 5 10 4 4 7 4 4 8 9 (шимпанзе)
11 12 11 11 10 8 10 8 12 10 12 12 12 11 11 11 (человек)
Это оказались еще более медленные маркеры, и 64 мутации (!) между ними помещают общего предка шимпанзе и человека на глубину времен в 4.4 миллионов лет назад. В целом, это же дают и антропологические оценки. Что важно — никаких «инопланетян» в создании современного человека не было, вместо этого имеется непрерывный эволюционный путь от общего предка шимпанзе и человека миллионы лет назад к нам, современным людям.
Можно ли считать, что все люди – родственники?
Как было отмечено в ответе на предыдущий вопрос, концептуально – да, но детали нам пока неизвестны. Если даже мы не найдем конкретного общего предка всех людей на Земле, как мужчин, так и женщин, но принципиальное сходство гаплотипов позволяет предполагать, что все люди произошли от одного общего предка. Если даже мы сможем найти нескольких, которых не сумеем свести к одному общему предку, все равно – опять интуитивно – он и она должны быть. В общем, дело опять сводится к определениям, как мы понимаем «общего предка».
Этот предок оказался древнее, чем предполагалось раньше. Еще недавно считалось, что он жил примерно 70 тыс. лет назад, потом 120 тысяч, потом 140 тысяч, сейчас он уже уходит глубже 200 тыс. лет назад, постепенно приближаясь к общему предку с неандертальцем 300-500 тыс. лет назад, и, возможно, уйдет еще далее к приматам, миллионы лет назад. Как мы видим, гаплотипы хоть и отдаленно, но в принципе похожи на те, что есть у всех нас. У шимпанзе и у нас — одни и те же маркеры. Следовательно, общий предок был один и тот же, только очень древний. И действительно, геномный анализ показывает, что шимпанзе из зоопарка и любой современный человек, мужчина или женщина, имеют более 95% совпадений в составе их ДНК, или в числе и положениях мутаций, как ни считать.
Насколько различаются гаплотипы у разных популяций? Как связать гаплотипы с субкладами?
Гаплотипы изображают в виде числа тандемов, или повторов, по каждому маркеру, выбранному из десятков и сотен. В англоязычной литературе их называют STR, или Short Tandem Repeats. Самый простой и короткий гаплотип из тех, которые рассматривает ДНК-генеалогия, состоит из пяти или шести маркеров.
Вот примеры – у этнических русских, славян с Русской равнины (индекс субклада R1a-Z280)
16 12 25 11 11 13
у скифов из раскопок в Минусинской котловине, с датировками 3800-3400 лет назад (индекс не определяли)
16 12 25 11 11 13
в высших кастах Индии (R1a-Z93-L342.2-L657) точно такая же, как у скифов
16 12 25 11 11 13
у современных таджиков гаплогруппы R1a
16 12 25 10 11 13 (хотя число 10 там при усреднении практически равно 10.5, и при добавлении гаплотипов может склониться или к 10, или к 11),
у носителей гаплогруппы R1a, наиболее распространенной у пуштунов Афганистана (индекс R1a-Z93-L342.2-Z2124)
16 12 24 11 11 13
у западных славян (индекс R1a-M458-L260)
17 12 25 10 11 13
Потомки древних носителей гаплогруппы R1a-M458 сейчас живут в основном в Белоруссии, Польше, на Украине, продвинулись в Центральную Европу; возможно, это они были первыми кельтами в Центральной Европе (в частности, в Австрии, образуя ядро гальштаттской археологической культуры). Как мы видим, 6-маркерный гаплотип уже другой, и легко отличается. Продолжим:
у славян «центрально-европейской группы» (R1a-M458-CTS11962)
16 12 25 10 11 13
у 60% западноевропейцев, носителей гаплогруппы R1b
14 12 24 11 13 13
Видно, что в ряде случаев у носителей той же гаплогруппы R1a даже короткие гаплотипы немного различаются, часто всего на одну мутацию, потому что они принадлежат разным субкладам, то есть племенам, с разной историей миграций, и с разными общими предками для каждого племени, хотя имеют одного общего предка гаплогруппы (R1a в данном случае). Поэтому надо всегда уточнять, о каком общем предке идет речь. Как уже говорилось выше, с расширением степени родства общий предок уходит в глубь времен, но он всегда есть.
Посмотрим, как это выглядит на сокращенной диаграмме субкладов гаплогруппы R1a c добавлением R1b:
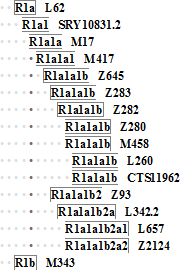
У каждой из этих гаплогрупп и субкладов был свой общий предок, и у каждого – свой гаплотип. Как мы уже увидели, 6-маркерные гаплотипы разделяют только тех общих предков, родоначальников, патриархов своих субкладов, у которых случайно проскочила мутация в тех самых 6-маркерных гаплотипах. Но у близко расположенных субкладов мутация – чисто статистически, неупорядоченно – могла и не проскочить. Так, мы уже видим, что восточные славяне Русской равнины (субклад R1a-Z280) – близкие родственники ариям (точнее, их потомкам из высших каст Индии) и скифам (ископаемые ДНК), у них одинаковые 6-маркерные гаплотипы. А гаплотипы пуштун, происходящих из исторической Бактрии, и западных славян, с которыми восточные славяне разделены десятью тысячами лет, неупорядоченные ошибки при копировании Y-хромосомы успели добавить за это время одну-две мутации. В гаплотипах R1a и R1b, которые разделены во времени десятками тысяч лет, уже есть пять мутаций разницы. Формальный расчет дает 5/0.0074 = 676 → 1580 условных поколений, то есть 40±18 тысяч лет расстояния между ними. Так оно в общем и получается, в пределах погрешности расчетов, между ними примерно 44 тысячи лет разницы.
Схема выше – предельно сокращенная, на самом деле в ней десятки уровней. Мы просто выделили кратчайший путь к ряду субкладов от образования исходной гаплогруппы R1a, которое произошло примерно 20 тысяч лет назад. От R1a последовательно отходили ветви новых и новых субкладов, в основе каждого была новая мутация в Y-хромосоме новорожденного мальчика, мужские потомки которого выжили и в итоге дали свое разветвленное потомство.
Примерно 5500 лет назад в Европе образовался так называемый юго-восточный субклад Z93, носители которого (потомки очередного мальчика, в Y-хромосоме которого образовалась мутация Z93) пришли на Русскую равнину, примерно 4800 лет назад там образовался очередной субклад L342.2/Z94, носители которого разошлись по нескольким направлениям. Одна часть продвинулась в Индию, с новообразованным субкладом L657, это были легендарные арии, и потомки индоариев в настоящее время продолжают нести в своих ДНК мутацию под индексом L657. Потому носителей L657 так много в высших кастах Индии, до 72% от общего количества. Другая часть продвинулась в Бактрию, но субклады таджиков пока малоизвестны. Тем не менее, датировка общих предков таджиков гаплогруппы R1a указывает на исторических ариев как их потоков. Еще одна часть ариев прошла в Иран и Афганистан, но у них был уже другой, «параллельный» субклад Z2124, которые сейчас имеют большинство пуштунов.
В итоге мы имеем две параллельные системы отсчета времен миграций и определения их направлений – гаплотипы (c их STR, маркерами, тандемными повторами блоков нуклеотидов) и субклады (с их SNP, снипами). Мутации в них происходят независимо друг от друга, но поскольку эти процессы идут параллельно, в одних и тех же популяциях, то в целом между мутационной динамикой в гаплотипах и субкладах есть некоторая корреляция. Мы уже видели, что и 6-маркерный гаплотип сдвинулся на одну мутацию между индусами и афганцами, носителями гаплогруппы R1a, и субклады сдвинулись тоже.
Мы видим, что даже короткие, 6-маркерные гаплотипы являются вполне информативными, и позволяют распознать представителей разных народов и (порой) этносов. Более протяженные гаплотипы, вплоть до 67- и 111-маркерных, позволяют уточнить картину, выявить более недавние ДНК-генеалогические линии, определить их датировку. Но суть поиска от этого принципиально не меняется, появляется намного более высокое разрешение в исследованиях.
Какова скорость мутаций в самых коротких, 6-маркерных гаплотипах?
Как уже пояснялось выше, гаплотипы показывают число повторов нуклеотидных участков в маркерах, которые в 6-маркерных гаплотипах обозначают соответствующими индексами 19, 388, 390, 391, 392 и 393. Например, у большинства афганцев гаплогруппы R1a в первом маркере (номер 393) было найдено 13 повторов определённой последовательности нуклеотидов. Во втором — 24 повтора уже другой последовательности и так далее. При передаче этого гаплотипа от отца сыну с вероятностью примерно 0.7% процента (то есть в среднем у одного на 135 рождений) может произойти мутация, например, такая – от гаплотипа
13-24-16-11-12-11
в такой:
13-25-16-11-12-11
А может произойти и такая –
13-24-15-11-12-11
Примеры таких мутаций можно найти в изобилии, тысячами. Может пройти и любая другая (как правило, одиночная, то есть на один шаг) мутация в любом маркере, только одни мутации в одних маркерах происходят чаще, в других — реже. А в среднем мутация в таких 6-маркерных гаплотипах происходит, как было найдено, в среднем раз на 135 мальчиков. Если перевести во времена, то константа скорости мутации в 6-маркерных гаплотипах равна 0.0074 мутаций на весь гаплотип за 25 лет.
Насколько обосновано положение, что по числу мутаций в гаплотипах можно определять «возраст гаплотипа», и не странно ли последнее понятие, поскольку речь о гаплотипах наших современников?
Действительно, «возраст гаплотипа» это жаргон, речь идет о времени, прошедшем со времени жизни общего предка рассматриваемой популяции. Иначе говоря, сколько времени прошло от предкового гаплотипа до гаплотипа его прямого потомка, нашего современника. То есть речь идет опять о времени до общего предка. Это и есть синоним жаргонного понятия «возраст гаплотипа».
Действительно, по числу мутаций в гаплотипах можно определять «возраст гаплотипа», то есть время, прошедшее от общего предка этого гаплотипа до его сегодняшних потомков. Поскольку сыновья в подавляющем числе случаев сохраняют гаплотип отца, переданный по наследству, и мутации в этом гаплотипе проскакивают в среднем только раз примерно в 3375 лет (6-маркерный гаплотип) или раз в 1250 лет (12-маркерный гаплотип), или раз в 125 лет (111-маркерный гаплотип), то даже через 5000 лет у потомков сохранится 23% исходного 6-маркерного гаплотипа, без изменений. То есть в списке из 100 гаплотипов потомков — 23 гаплотипа будут такими же, какой был у предка 5000 лет назад. Это несложно проверить, применяя логарифмический метод [ln(100/23)]/0.0074 = 199 условных поколений (по 25 лет), то есть примерно 5000 лет. На самом деле 23 гаплотипа из ста сохранятся заметно дольше, чем пять тысяч лет, потому что в гаплотипах время от времени происходят возвратные мутации, как бы «стирающие» мутации, образовавшиеся к тому времени. Расчет показывается, что 23 гаплотипа (6-маркерных) из ста сохранятся через 199 → 227 условных поколений, то есть через 5675 лет. Здесь стрелка показывает поправку на возвратные мутации.
При рассмотрении 12-маркерных гаплотипов те же 23% гаплотипов предка сохранятся через 73 поколения (без поправки на возвратные мутации), или 78 поколений, то есть 1950 лет. Таким образом, определить гаплотип предка можно и через тысячелетия. И по его виду можно узнать, из каких краёв предок пришёл, сравнив вид гаплотипа с гаплотипами по территориям, и с доступными ископаемыми гаплотипами. В качестве примера стабильности гаплотипов можно привести 25-маркерные гаплотипы дяди и племянника (гаплогруппа R1b-U152), которые оказались идентичными (это гаплотипы KLIN00012 и KLIN00013 в базе данных Академии ДНК-генеалогии):
13 25 15 11 11 11 12 12 12 12 13 28 – 19 9 10 11 11 25 14 19 29 15 15 16 17
Как видно, за три поколения, разделяющих дядю и племянника, 25-маркерный гаплотип полностью сохранился. Действительно, расчеты показывают, что в среднем одна мутация между двумя 25-маркерными гаплотипами случается только на протяжении 11 поколений: 1/2/0.046 = 10.87 условных поколений (по 25 лет каждое; 0.046 – константа скорости мутации в 25-маркерных гаплотипах). Можно было бы написать «в среднем через 11 поколений», но это было бы по сути неверно, так как мутация может произойти неупорядоченно и когда угодно, но при наличии большого числа опытов она происходит в среднем раз в 11 поколений.
Можно ли по гаплотипам определять этносы?
Как правило, нет. Гаплотипы не указывают на этносы, это совершенно разные понятия. Гаплотипы указывают на древние рода, племена, которые намного старше этносов. С тех древних времен гаплотипы давно разошлись по разным территориям, на которых потом, через тысячелетия сформировались этносы. Хотя нередко бывают ситуации, когда относительно молодой этнос имеет характерный гаплотип, который мутировал всего лишь незначительно (то есть относительно недавно) от общего предка, и легко узнается. Например, таким характерным является гаплотип евреев гаплогруппы R1a. Он вошел в еврейскую среду субкладом Z2124 (тем самым, который мы видим у афганцев) примерно 4000 лет назад, но у афганцев этот субклад появился только в начале нашей эры. 1300 лет назад группа евреев, носителей этого субклада, к которому за прошедшие тысячелетия добавилась мутация М582 (и цепочка субкладов евреев приобрела вид R1a-Z93-L342.2-Z2124-M582), по каким-то причинам почти полностью вымерла, видимо, была группой компактной, и выживший носитель мутации М582 фактически опять начал свой род. Поскольку это было всего 1300 лет назад, то гаплотип в значительной степени сохранился до настоящего времени, и сейчас почти у всех евреев гаплогруппы R1a, а их многие тысячи, гаплотип (в 67-маркерном формате) имеет вид:
13 25 16 10 11 14 12 12 10 13 11 30 — 14 9 11 11 11 24 14 20 30 12 12 15 15 — 11 11 19 23 14 16 19 20 35 38 14 11 — 11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 14 8 14 23 21 12 12 11 13 10 11 12 13
В среднем, у всех евреев, носителей этого гаплотипа, наблюдается всего 6 мутаций (на 67 маркерах) от предкового гаплотипа с возрастом 1300 лет. Поэтому данный гаплотип распознается с одного взгляда опытного специалиста. В нем есть характерные только для данного гаплотипа фрагменты.
У афганцев есть тоже характерные по виду гаплотипы. Например, в гаплогруппе G2 в Афганистане преобладает субклад G2b1, c коротким 6-маркерным гаплотипом
13 23 16 11 12 11
а на Кавказе преобладает субклад G2a1, с гаплотипом
14 22 15 10 12 10
Здесь различия множественные, поскольку общий предок обоих субкладов жил не менее 15 тысяч лет назад. За это время гаплотипы разошлись столь далеко друг от друга. Поскольку у каждого племени и в каждом этносе, в каждом регионе можно определять возраст племени и вообще популяции в целом, то можно определять, когда и в каком направлении шли древние миграции.
Можно ли сказать, что где определенной гаплогруппы или субклада больше всех, там и их прародина?
Нет, так сказать нельзя, во всяком случае, в большинстве случаев. Например, в Ирландии гаплогруппы R1b больше 90%, но гаплогруппа R1b появилась наиболее вероятно в Южной Сибири, причем примерно 20 тысяч лет назад, а «возраст» гаплогруппы R1b в Ирландии, как и вообще в Европе, менее 5 тысяч лет. Причина – в далеких древних миграциях. Носители гаплогруппы R1b прибыли в Европу в начале III тыс. до н.э., причем основная миграция была со стороны Пиреней, и оттуда мигранты довольно быстро заселили Британские острова, и далее распространидись по Европе в виде культуры колоколовидных кубков. К настоящему времени уже изучили шесть ископаемых ДНК из культуры колоколовидных кубков, и из них пять оказались R1b, и один – R1, для всех датировки между 4500 и 4100 лет назад.
В периферийных регионах Европы (Пиренейский полуостров, Британские острова) потомки мигрантов быстро приумножили свое количество, и сейчас доля гаплогруппы R1b с нисходящими субкладами составляет там около 90% или выше, а в центральной Европе произошло значительное разбавление новыми мигрантами, уже других гаплогрупп. В Восточной Европе исторически сложилось так, что там около 50% составляет гаплогруппа R1a, но там тоже не их прародина, несмотря на высокую долю гаплогруппы. В Финляндии – выше 60% гаплогруппы N, но это тоже не ее прародина. В Финляндию носители гаплогруппы N пришли со стороны Алтая. В Осетии более 70% гаплогруппы G2a, но и это не прародина. Состав и доля гаплогрупп складывается тысячелетиями, и отражает определенные исторические процессы, о которых историки во многих случаях и не догадываются.
Полагать, что там, где гаплогруппы больше, там и прародина, это застарелая ошибка популяционных генетиков. С этой ошибки фактически и началась популяционная генетика человека, когда решили, что высокое содержание гаплогруппы R1b в Европе указывает на то, что эта гаплогруппа в Европе и образовалась. Более того, в академической литературе появилась и датировка для образования R1b в Европе – 30 тысяч лет назад. Эту датировку попгенетики тут же стали обильно цитировать, и никто не задавал вопрос – как эту величину определили? На самом деле ее никто не определял, она была просто записана «по понятиям». Это – типичная схема для популяционных генетиков.
Вывод – доля в процентах гаплогруппы в популяции сама по себе означает не очень много, это просто структура современной популяции, то, чем занимаются популяционные генетики. Часто бывает, что доля большая, а общий предок недавний, просто потомки общего предка быстро размножились, условия были благоприятными. Еще пример – доля гаплогруппы R1a в высших кастах Индии достигает 72%, а сама гаплогруппа пришла в Индию с ариями примерно 3500 лет назад. Хотя в самой цитируемой работе по той же порочной «логике» попгенетиков утверждалось, что раз доля гаплогруппы R1a в Индии (то есть в высших кастах) столь высока, то R1a в Индии и образовалась. На что можно с иронией сказать, что поскольку в нашей фамильной деревне Клёсово в Курской области доля гаплогруппы R1a составляет 100%, то гаплогруппа в той деревне и образовалась. Поскольку у каждого племени и в каждом этносе, в каждом регионе можно определять возраст племени и вообще популяции в целом, то можно определять, когда и в каком направлении шли древние миграции.
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке

















